A Meta-Prediction of Methylenetetrahydrofolate-Reductase Polymorphisms and Air Pollution Increased the Risk of Ischemic Heart Diseases Worldwide


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Selection and Data Extraction
2.2. Quality Assessment
2.3. Data Synthesis and Analysis
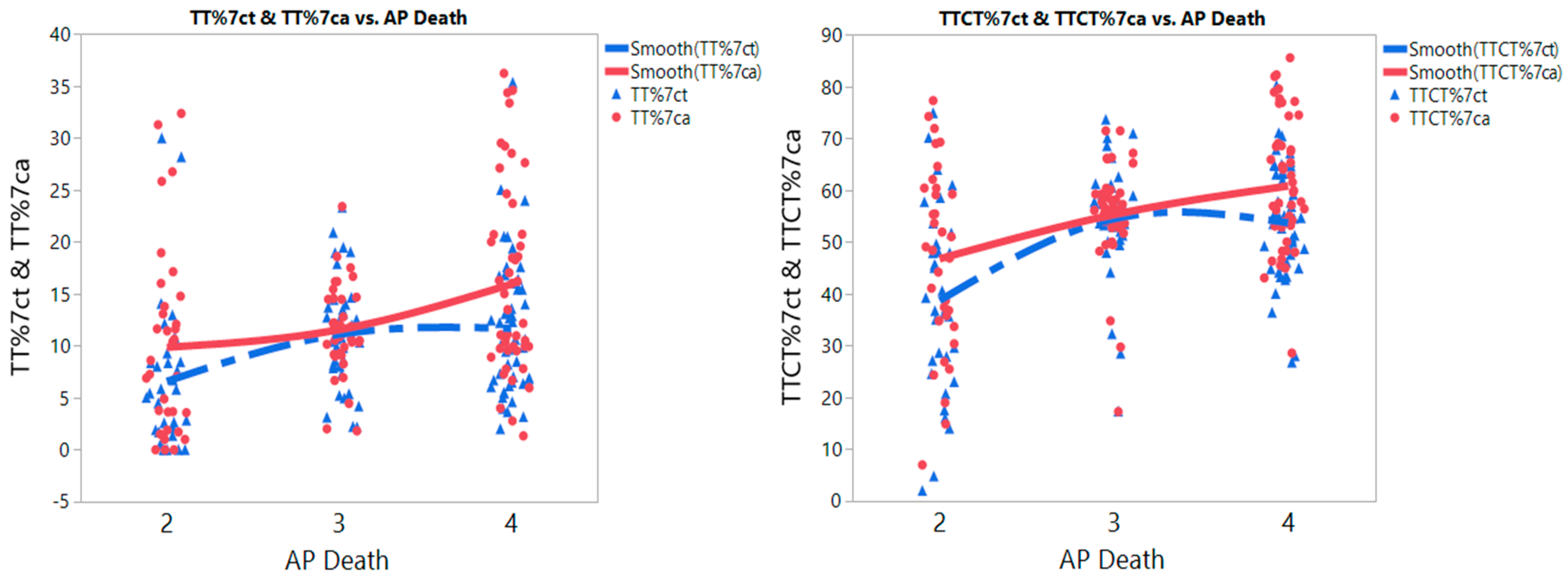
3. Results
3.1. Sample Characteristics and Genotype Frequency
3.2. Pooled Meta-Analysis
3.3. Subgroup Analysis by Ethnicity and Countries
3.4. Subgroup Analysis by IHD Subtypes
3.5. Heterogeneous Findings by GIS Map
3.6. Meta-Prediction: MTHFR Polymorphisms and Air Pollution Associated with Risk of IHD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Executive summary: Heart disease and stroke statistics—2014 update: A report from the American Heart Association. Circulation 2014, 129, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Nichols, M.; Townsend, N.; Scarborough, P.; Rayner, M. Cardiovascular disease in Europe 2014: Epidemiological update. Eur. Heart J. 2014, 35, 2950–2959. [Google Scholar] [CrossRef] [PubMed]
- Nabel, E.G.; Braunwald, E. A tale of coronary artery disease and myocardial infarction. N. Engl. J. Med. 2012, 366, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart disease and stroke statistics—2017 update: A report from the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Finegold, J.A.; Asaria, P.; Francis, D.P. Mortality from ischaemic heart disease by country, region, and age: Statistics from World Health Organisation and United Nations. Int. J. Cardiol. 2013, 168, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, I.I.; Tillmann, T.; Banerjee, A. Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: A systematic analysis for the global burden of disease study 2013. Lancet 2015, 385, 117–171. [Google Scholar] [CrossRef]
- Peng, H.Y.; Man, C.F.; Xu, J.; Fan, Y. Elevated homocysteine levels and risk of cardiovascular and all-cause mortality: A meta-analysis of prospective studies. J. Zhejiang Univ. Sci. B 2015, 16, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Je, Y. Dietary fiber intake and total mortality: A meta-analysis of prospective cohort studies. Am. J. Epidemiol. 2014, 180, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, X.; Liu, J.; Xie, X.; Cui, W.; Zhu, Y. Homocysteine accelerates senescence of endothelial cells via DNA hypomethylation of human telomerase reverse transcriptase. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Kones, R. Molecular sources of residual cardiovascular risk, clinical signals, and innovative solutions: Relationship with subclinical disease, undertreatment, and poor adherence: Implications of new evidence upon optimizing cardiovascular patient outcomes. Vasc. Health Risk Manag. 2013, 9, 617–670. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.M.; Dear, A.; Craig, J.M.; Norman, P.E.; Golledge, J. The potential role of homocysteine mediated DNA methylation and associated epigenetic changes in abdominal aortic aneurysm formation. Atheroscler. 2013, 228, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Gil Nunez, A.C. Homocysteine: Vascular risk factor? Neurologia 2004, 19, 1–4. [Google Scholar] [PubMed]
- Luo, H.; Liu, B.; Hu, J.; Wang, X.; Zhan, S.; Kong, W. Hyperhomocysteinemia and methylenetetrahydrofolate reductase polymorphism in cervical artery dissection: A meta-analysis. Cerebrovasc. Dis. 2014, 37, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Hasan, N.; Marjot, T.; Khan, M.S.; Prasad, K.; Bentley, P.; Sharma, P. Detailed analysis of gene polymorphisms associated with ischemic stroke in south Asians. PLoS ONE 2013, 8, e57305. [Google Scholar] [CrossRef] [PubMed]
- Klerk, M.; Verhoef, P.; Clarke, R.; Blom, H.J.; Kok, F.J.; Schouten, E.G. Mthfr 677c→t polymorphism and risk of coronary heart disease: A meta-analysis. JAMA 2002, 288, 2023–2031. [Google Scholar] [CrossRef] [PubMed]
- Langrish, J.P.; Li, X.; Wang, S.; Lee, M.M.; Barnes, G.D.; Miller, M.R.; Cassee, F.R.; Boon, N.A.; Donaldson, K.; Li, J.; et al. Reducing personal exposure to particulate air pollution improves cardiovascular health in patients with coronary heart disease. Environ. Health Perspect. 2012, 120, 367–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, W.Q.; Koehoorn, M.; Davies, H.W.; Demers, P.A.; Tamburic, L.; Brauer, M. Long-term exposure to traffic-related air pollution and the risk of coronary heart disease hospitalization and mortality. Environ. Health Perspect. 2011, 119, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Brook, R.D.; Rajagopalan, S.; Pope, C.A., 3rd; Brook, J.R.; Bhatnagar, A.; Diez-Roux, A.V.; Holguin, F.; Hong, Y.; Luepker, R.V.; Mittleman, M.A.; et al. Particulate matter air pollution and cardiovascular disease: An update to the scientific statement from the American Heart Association. Circulation 2010, 121, 2331–2378. [Google Scholar] [CrossRef] [PubMed]
- Brook, R.D.; Franklin, B.; Cascio, W.; Hong, Y.; Howard, G.; Lipsett, M.; Luepker, R.; Mittleman, M.; Samet, J.; Smith, S.C.; et al. Air pollution and cardiovascular disease: A statement for healthcare professionals from the expert panel on population and prevention science of the American Heart Association. Circulation 2004, 109, 2655–2671. [Google Scholar] [CrossRef] [PubMed]
- Dominici, F.; Peng, R.D.; Bell, M.L.; Pham, L.; McDermott, A.; Zeger, S.L.; Samet, J.M. Fine particulate air pollution and hospital admission for cardiovascular and respiratory diseases. JAMA 2006, 295, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Ruckerl, R.; Ibald-Mulli, A.; Koenig, W.; Schneider, A.; Woelke, G.; Cyrys, J.; Heinrich, J.; Marder, V.; Frampton, M.; Wichmann, H.E.; et al. Air pollution and markers of inflammation and coagulation in patients with coronary heart disease. Am. J. Respir. Crit. Care Med. 2006, 173, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Dockery, D.W.; Muller, J.E.; Mittleman, M.A. Increased particulate air pollution and the triggering of myocardial infarction. Circulation 2001, 103, 2810–2815. [Google Scholar] [CrossRef] [PubMed]
- Van Meurs, J.B.; Pare, G.; Schwartz, S.M.; Hazra, A.; Tanaka, T.; Vermeulen, S.H.; Cotlarciuc, I.; Yuan, X.; Mälarstig, A.; Bandinelli, S.; et al. Common genetic loci influencing plasma homocysteine concentrations and their effect on risk of coronary artery disease. Am. J. Clin. Nutr. 2013, 98, 668–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wald, D.S.; Morris, J.K.; Wald, N.J. Reconciling the evidence on serum homocysteine and ischaemic heart disease: A meta-analysis. PLoS ONE 2011, 6, e16473. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Bennett, D.A.; Parish, S.; Verhoef, P.; Dötsch-Klerk, M.; Lathrop, M.; Xu, P.; Nordestgaard, B.G.; Holm, H.; Hopewell, J.C.; et al. Homocysteine and coronary heart disease: Meta-analysis of MTHFR case-control studies, avoiding publication bias. PLoS Med. 2012, 9, e1001177. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Hong, K.; Hu, J.; Cheng, X. Three genetic polymorphisms of homocysteine-metabolizing enzymes and risk of coronary heart disease: A meta-analysis based on 23 case-control studies. DNA Cell Biol. 2012, 31, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Jee, S.H.; Beaty, T.H.; Suh, I.; Yoon, Y.; Appel, L.J. The methylenetetrahydrofolate reductase gene is associated with increased cardiovascular risk in Japan, but not in other populations. Atherosclerosis 2000, 153, 161–168. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The prisma statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Sagoo, G.S.; Little, J.; Higgins, J.P. Systematic reviews of genetic association studies. Human genome epidemiology network. PLoS Med. 2009, 6, e28. [Google Scholar] [CrossRef] [PubMed]
- Girelli, D.; Friso, S.; Trabetti, E.; Olivieri, O.; Russo, C.; Pessotto, R.; Faccini, G.; Pignatti, P.F.; Mazzucco, A.; Corrocher, R. Methylenetetrahydrofolate reductase C677T mutation, plasma homocysteine, and folate in subjects from northern Italy with or without angiographically documented severe coronary atherosclerotic disease: Evidence for an important genetic-environmental interaction. Blood 1998, 91, 4158–4163. [Google Scholar] [PubMed]
- Morita, H.; Taguchi, J.I.; Kurihara, H.; Kitaoka, M.; Kaneda, H.; Kurihara, Y.; Maemura, K.; Minamino, T.; Ohno, M.; Yamaoki, K.; et al. Genetic polymorphism of 5,10-methylenetetrahydrofolate reductase (MTHFR) as a risk factor for coronary artery disease. Circulation 1997, 95, 2032–2036. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.P.K.; Yu, C.H. Meta-prediction of MTHFR gene polymorphism mutations and associated risk for colorectal cancer. Biol. Res. Nurs. 2016, 18, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Deeks, J.J.; Higgins, J.; Altman, D.G. Analysing data and undertaking meta-analyses. In Cochrane Handbook for Systematic Reviews of Interventions: Cochrane Book Series; Green, S., Higgins, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2008; pp. 243–296. [Google Scholar]
- Duval, S.; Tweedie, R. Trim and fill: A simple funnel-plot-based method of testing and adjusting for publication bias in meta-analysis. Biometrics 2000, 56, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Davey Smith, G.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speybroeck, N. Classification and regression trees. Int. J. Pub. Health 2012, 57, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, J.; Becker, M.A.; Wood, G. Pairwise multiple comparison procedures: A review. Psychol. Bull. 1984, 96, 589. [Google Scholar] [CrossRef]
- Albrecht, J. Key Concepts and Techniques in GIS; Sage: Thousand Oaks, CA, USA, 2007. [Google Scholar]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Chapman and Hall/CRC: Boca Raton, FL, USA, 1984. [Google Scholar]
- Bala, M.; Agrawal, R.K. Statistical measures to determine optimal structure of decision tree: One versus one support vector machine. Def. Sci. J. 2010, 60, 399–404. [Google Scholar] [CrossRef]
- Akaike, H. Akaike's Information Criterion; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In International Symposium on Information Theory; Petrov, B., Csaki, F., Eds.; Akademia Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Symonds, M.R.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Kenworthy, J.; Laube, F. Urban transport patterns in a global sample of cities & their linkages to transport infrastructure, land use, economics & environment. World Transp. Policy Prac. 2002, 8, 5–20. [Google Scholar]
- World Health Organization. The Urban Environment. 2015. Available online: http://www.who.int/heli/risks/urban/urbanenv/en/ (accessed on 27 February 2018).
- US Environmental Protection Agency. Air Quality Index Basics. Available online: https://www.airnow.gov/index.cfm?action=aqibasics.aqi (accessed on 2 January 2018).
- World Health Organization. Deaths Attributable to Urban Air Pollution. 2004. Available online: http://www.who.int/heli/risks/urban/en/uapmap.1.pdf?ua¼1 (accessed on 2 January 2018).
- World Health Organization. Global Health Risks. 2009. Available online: http://www.who.int/healthinfo/global_burden_disease/GlobalHealthRisks_report_full.pdf (accessed on 2 January 2018).
- World Health Organization. Global Health Risks. 2012. Available online: https://commons.wikimedia.org/wiki/File:Deaths_from_air_pollution.png (accessed on 2 January 2018).
- American Institute for Cancer Research. How AICR Recommendations Cuts Colorectal Cancer Risk for Both Men and Women. Available online: http://www.aicr.org/cancer-research-update/2016/11_02/cru-how-AICR-recommendations-cuts-colorectal-cancer-risk-for-men-and-women.html (accessed on 16 March 2018).
- Viera, A.J. Odds ratios and risk ratios: What’s the difference and why does it matter? South Med. J. 2008, 101, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Scott, I. Interpreting risks and ratios in therapy trials. Aust. Prescr. 2008, 31, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Team, Q.D.; Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project. 2012. Available online: http://qgis.osgeo.org (accessed on 2 January 2018).
- Bentley, P.; Peck, G.; Smeeth, L.; Whittaker, J.; Sharma, P. Causal relationship of susceptibility genes to ischemic stroke: Comparison to ischemic heart disease and biochemical determinants. PLoS ONE 2010, 5, e9136. [Google Scholar] [CrossRef] [PubMed]
- Drenos, F.; Whittaker, J.C.; Humphries, S.E. The use of meta-analysis risk estimates for candidate genes in combination to predict coronary heart disease risk. Ann. Hum. Genet. 2007, 71, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Chen, X.; Shi, J. Genetic polymorphism of MTHFR C677T and premature coronary artery disease susceptibility: A meta-analysis. Gene 2015, 565, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Yang, H.L.; Shiao, S.P. Meta-prediction of MTHFR gene polymorphisms and air pollution on the risk of hypertensive disorders in pregnancy worldwide. Int. J. Environ. Res. Public Health 2018, 15, E326. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.M.; Chen, Z.F.; Young, L.; Shiao, S.P.K. Meta-prediction of the effects of methylenetetrahydrofolate reductase gene polymorphisms and air pollution on risk of Alzheimer’s disease. Int. J. Environ. Res. Public Health 2017, 14, E63. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, M.C.; Yu, P.J.; Shiao, S.P.K. Meta-prediction of MTHFR gene polymorphism-mutations and air pollution as risk factors for breast cancer. Nurs. Res. 2017, 66, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Lien, S.Y.A.; Young, L.; Gau, B.S.; Shiao, S.P.K. Meta-prediction of MTHFR gene polymorphism-mutations, air pollution, and risks of leukemia among world populations. Oncotarget 2017, 8, 4387–4398. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fang, T.B. Examining personal air pollution exposure, intake, and health danger zone using time geography and 3D geovisualization. ISPRS Int. J. Geo-Inf. 2015, 4, 32–46. [Google Scholar] [CrossRef]
- Leclerc, D.; Sibani, S.; Rozen, R. Molecular Biology of Methylenetetrahydrofolate Reductase (MTHFR) and Overview of Mutations/Polymorphisms. In MTHFR Polymorphisms and Disease; CRC Press: Boca Raton, FL, USA, 2005; pp. 15–34. [Google Scholar]
- Lee, B.J.; Kim, B.; Lee, K. Air pollution exposure and cardiovascular disease. Toxicol. Res. 2014, 30, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Packard, G. Review article: On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Biol. J. Linn. Soc. 2014, 113, 1167–1178. [Google Scholar] [CrossRef]
- Xiao, X.; White, E.P.; Hooten, M.B.; Durham, S.L. On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Ecology 2011, 92, 1887–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.H.; Lee, H.S.; Gan, S.; Brown, E. Nonlinear modeling with big data in SAS and JMP. In Proceedings of the Western Users of SAS Software Conference, Long Beach, CA, USA, 20–22 September 2017. [Google Scholar]
- Choi, I.H.; Noh, J.S.; Han, J.S.; Kim, H.J.; Han, E.S.; Song, Y.O. Kimchi, a fermented vegetable, improves serum lipid profiles in healthy young adults: Randomized clinical trial. J. Med. Food. 2013, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Lista, J.; Perez-Martinez, P.; Garcia-Rios, A.; Alcala-Diaz, J.F.; Perez-Caballero, A.I.; Gomez-Delgado, F.; Fuentes, F.; Quintana-Navarro, G.; Lopez-Segura, F.; Ortiz-Morales, A.M.; et al. CORonary Diet Intervention with Olive oil and cardiovascular PREVention study (the CORDIOPREV study): Rationale, methods, and baseline characteristics: A clinical trial comparing the efficacy of a Mediterranean diet rich in olive oil versus a low-fat diet on cardiovascular disease in coronary patients. Am. Heart J. 2016, 177, 42–50. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MTHFR 677 | IHD | CAD | MI |
|---|---|---|---|
| (n Case/n Control) | (n Case/n Control) | (n Case/n Control) | |
| 123 Studies (29,697/31,028) | 93 Studies (22,994/20,221) | 30 Studies (6709/10,807) | |
| Overall | Risk Type: TT, CT | Risk Type: TT, CT | |
| Protective: CC | Protective: CC | Protective: CC | |
| Subgroups | |||
| Caucasian | 60 Studies | 47 Studies | 13 Studies |
| (22,042/14,925) | (13,817/10,702) | (2850/4223) | |
| Risk trend: TT | Risk Type: TT | NS | |
| Protective trend: CC | Protective: CC | ||
| East Asian | 32 Studies | 25 Studies | 7 Studies |
| (7112/9471) | (4855/4931) | (2257/4540) | |
| Risk Type: TT | Risk Type: TT | Risk Type: TT | |
| Protective: CC | Protective: CC | ||
| South Asian | 13 Studies | 8 Studies | 5 Studies |
| (2475/2763) | (1447/1621) | (1028/1142) | |
| Risk Type: TT, CT | Risk Type: TT, CT | NS | |
| Protective: CC | Protective: CC | ||
| Mixed group | 3 Studies | 2 Studies | 1 Study |
| (951/852) | (882/514) | (69/338) | |
| NS | NS | -- | |
| Middle Eastern | 7 Studies | 6 Studies | 1 Study |
| (1265/1723) | (1169/1623) | (99/100) | |
| Risk Type: TT, CT | Risk Type: TT, CT | -- | |
| Protective: CC | Protective: CC | ||
| Hispanic | 2 Studies | -- | 2 Studies |
| (353/364) | (353/364) | ||
| NS | NS | ||
| African | 6 Studies | 5 Studies | 1 Study |
| (874/930) | (824/648) | (50/282) | |
| Risk Type: TT | Risk Type: TT, CT | -- |
| Genotype per Ethnicity (n Studies) | IHD Cases (n = 29,697) n (%) | Controls (n = 31,028) n (%) | Test of Heterogeneity | Test of Association | |||||
|---|---|---|---|---|---|---|---|---|---|
| Q | p * | I2 | Risk Ratio (95% CI) | p | |||||
| TT (123) | 3812 | (12.84) | 3330 | (10.73) | 238.56 | <0.0001 | 48.9% | 1.23 [1.14, 1.33] | <0.0001 |
| Caucasian (60) | 1850 | (11.10) | 1479 | (9.91) | 59.38 | 0.4615 | 0.6% | 1.07 [1.00, 1.14] | 0.0528 |
| East Asian (32) | 1439 | (20.23) | 1443 | (15.24) | 60.75 | 0.0011 | 49% | 1.31 [1.17, 1.47] | <0.0001 |
| South Asian (13) | 93 | (3.76) | 71 | (2.57) | 20.19 | 0.0635 | 40.6% | 1.51 [1.13, 2.01] | 0.0048 |
| Mixed Group (3) | 93 | (9.78) | 88 | (10.33) | 0.05 | 0.9771 | 0% | 0.86 [0.63, 1.17] | 0.33 |
| Middle Eastern (7) | 130 | (10.28) | 89 | (5.17) | 27.38 | 0.0001 | 78.1% | 2.62 [1.37, 5.01] | 0.0034 |
| Hispanic (2) | 102 | (28.90) | 106 | (29.12) | 1.50 | 0.2204 | 33.4% | 0.99 [0.79, 1.25] | 0.94 |
| African (6) | 105 | (12.01) | 54 | (5.81) | 2.03 | 0.8454 | 0% | 2.14 [1.56, 2.94] | <0.0001 |
| CT (123) | 12,777 | (43.02) | 12,815 | (41.30) | 174.49 | 0.0013 | 30.1% | 1.04 [1.01, 1.07] | 0.0028 |
| Caucasian (60) | 7402 | (44.41) | 6448 | (43.20) | 47.63 | 0.8555 | 0% | 1.01 [0.98, 1.04] | 0.48 |
| East Asian (32) | 3338 | (46.93) | 4439 | (46.87) | 53.81 | 0.0067 | 42.4% | 1.03 [0.98, 1.09] | 0.27 |
| South Asian (13) | 634 | (25.62) | 531 | (19.22) | 27.60 | 0.0063 | 56.5% | 1.32 [1.12, 1.57] | 0.0011 |
| Mixed Group (3) | 415 | (43.64) | 327 | (38.38) | 1.44 | 0.4874 | 0% | 1.02 [0.90, 1.15] | 0.78 |
| Middle Eastern (7) | 489 | (38.66) | 593 | (34.42) | 5.67 | 0.4607 | 0% | 1.18 [1.07, 1.30] | 0.0011 |
| Hispanic (2) | 165 | (46.74) | 157 | (43.13) | 1.88 | 0.1709 | 46.7% | 1.08 [0.92, 1.27] | 0.34 |
| African (6) | 334 | (38.22) | 320 | (34.41) | 9.31 | 0.0975 | 46.3% | 1.13 [1.00, 1.28] | 0.0547 |
| CC (123) | 13,108 | (44.14) | 14,883 | (47.97) | 269.85 | <0.0001 | 54.8% | 0.91 [0.89, 0.94] | <0.0001 |
| Caucasian (60) | 7415 | (44.49) | 6998 | (46.89) | 63.45 | 0.3225 | 7% | 0.98 [0.95, 1.00] | 0.058 |
| East Asian (32) | 2335 | (32.83) | 3589 | (37.89) | 89.99 | <0.0001 | 65.6% | 0.84 [0.77, 0.91] | <0.0001 |
| South Asian (13) | 1748 | (70.63) | 2161 | (78.21) | 44.14 | <0.0001 | 72.8% | 0.91 [0.86, 0.96] | 0.0011 |
| Mixed Group (3) | 443 | (46.58) | 437 | (51.29) | 1.09 | 0.5804 | 0% | 1.02 [0.92, 1.12] | 0.76 |
| Middle Eastern (7) | 646 | (51.07) | 1041 | (60.42) | 16.22 | 0.0126 | 63% | 0.76 [0.66, 0.87] | <0.0001 |
| Hispanic (2) | 86 | (24.36) | 101 | (27.75) | 0.04 | 0.8487 | 0% | 0.88 [0.69, 1.13] | 0.31 |
| African (6) | 435 | (49.77) | 556 | (59.78) | 11.88 | 0.0365 | 57.9% | 0.87 [0.76, 1.00] | 0.0501 |
| TT + CT (123) | 16,589 | (55.86) | 16,145 | (52.03) | 283.27 | <0.0001 | 56.9% | 1.09 [1.06, 1.11] | <0.0001 |
| CC + CT (123) | 25,885 | (87.16) | 27,698 | (89.27) | 304.35 | <0.0001 | 59.9% | 0.98 [0.97, 0.99] | <0.0001 |
| T (123) | 10,200 | (34.35) | 9737 | (31.38) | 180.11 | 0.0005 | 32.3% | 1.11 [1.07, 1.14] | <0.0001 |
| C (123) | 19,496 | (65.65) | 21,290 | (68.62) | 172.23 | 0.0019 | 29.2% | 0.96 [0.94, 0.97] | <0.0001 |
| Subgroups | |||||||||
| TT Risk > 1 (87) 1 | 18,032 | (60.72) | 22,413 | (72.23) | |||||
| TT | 2515 | (13.95) | 2351 | (10.49) | 163.42 | <0.0001 | 47.4% | 1.40 [1.28, 1.53] | <0.0001 |
| CT | 7720 | (42.81) | 9243 | (41.24) | 148.57 | <0.0001 | 42.1% | 1.05 [1.02, 1.09] | 0.0029 |
| CC | 7797 | (43.24) | 10,819 | (48.27) | 208.20 | <0.0001 | 58.7% | 0.88 [0.85, 0.91] | <0.0001 |
| TT + CT | 10,235 | (56.76) | 11,594 | (51.73) | 225.28 | <0.0001 | 61.8% | 1.13 [1.09, 1.17] | <0.0001 |
| CC + CT | 15,517 | (86.05) | 20,062 | (89.51) | 267.73 | <0.0001 | 67.9% | 0.96 [0.95, 0.98] | <0.0001 |
| T | 6375 | (35.35) | 6972 | (31.11) | 134.06 | 0.0007 | 35.8% | 1.17 [1.12, 1.22] | <0.0001 |
| C | 11,657 | (64.65) | 15,440 | (68.89) | 128.35 | 0.0021 | 33% | 0.94 [0.92, 0.95] | <0.0001 |
| TT Risk < 1 (12) 2 | 2731 | (9.20) | 2156 | (6.95) | |||||
| TT | 322 | (11.79) | 299 | (13.87) | 7.89 | 0.7229 | 0% | 0.86 [0.75, 1.00] | 0.051 |
| CT | 1151 | (42.15) | 848 | (39.33) | 8.71 | 0.6489 | 0% | 1.04 [0.97, 1.12] | 0.23 |
| CC | 1258 | (46.06) | 1009 | (46.80) | 11.75 | 0.3828 | 6.4% | 1.00 [0.95, 1.06] | 0.90 |
| TT + CT | 1473 | (53.94) | 1147 | (53.20) | 11.24 | 0.4235 | 2.1% | 1.00 [0.95, 1.05] | 0.90 |
| CC + CT | 2409 | (88.21) | 1857 | (86.13) | 13.30 | 0.2745 | 17.3% | 1.02 [1.00, 1.04] | 0.052 |
| T | 897.5 | (32.86) | 723 | (33.53) | 6.66 | 0.8262 | 0% | 0.97 [0.90, 1.05] | 0.44 |
| C | 1833.5 | (67.14) | 1433 | (66.47) | 7.30 | 0.7742 | 0% | 1.02 [0.98, 1.06] | 0.44 |
| TT Risk~1 (24) 3 | 8934 | (30.08) | 6459 | (20.82) | |||||
| TT | 975 | (10.91) | 680 | (10.53) | 18.02 | 0.7562 | 0% | 0.99 [0.90, 1.09] | 0.86 |
| CT | 3906 | (43.72) | 2724 | (42.17) | 16.19 | 0.8468 | 0% | 1.01 [0.97, 1.05] | 0.73 |
| CC | 4053 | (45.37) | 3055 | (47.30) | 17.43 | 0.7875 | 0% | 1.00 [0.96, 1.03] | 0.81 |
| TT + CT | 4881 | (54.63) | 3404 | (52.70) | 16.51 | 0.8325 | 0% | 1.00 [0.97, 1.04] | 0.81 |
| CC + CT | 7959 | (89.09) | 5779 | (89.47) | 18.51 | 0.7295 | 0% | 1.00 [0.99, 1.01] | 0.86 |
| T | 2928 | (32.77) | 2042 | (31.61) | 8.89 | 0.9963 | 0% | 1.00 [0.95, 1.05] | 0.95 |
| C | 6006 | (67.23) | 4417 | (68.39) | 9.19 | 0.9953 | 0% | 1.00 [0.98, 1.02] | 0.95 |
| Variable | Partition Tree | Tukey’s Test | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AICc | AP Death Levels | Count | Mean | SD | Levels Compared | Difference | SE Difference | Lower CI | Upper CI | p | |
| TT%ct | 802 | 2 | 34 | 6.6 | 7.0 | 4/2 | 5.1 | 1.4 | 1.8 | 8.4 | 0.0011 |
| 3 and 4 | 89 | 11 | 6.0 | 3/2 | 4.5 | 1.5 | 1.1 | 8.0 | 0.0064 | ||
| 4/3 | 0.6 | 1.3 | −2.6 | 3.7 | 0.91 | ||||||
| TT%ca | 857 | 2 and 3 | 74 | 11 | 6.8 | 4/2 | 6.1 | 1.7 | 1.9 | 10 | 0.0019 |
| 4 | 49 | 16 | 9.0 | 4/3 | 4.4 | 1.7 | 0.5 | 9.3 | 0.0251 | ||
| 3/2 | 1.7 | 1.8 | −2.6 | 6.0 | 0.63 | ||||||
| CT%ct | 894 | 2 | 34 | 32 | 13 | 3/2 | 11 | 2.1 | 6.3 | 16 | <0.0001 |
| 3 and 4 | 89 | 43 | 7.0 | 4/2 | 9.9 | 2.0 | 5.1 | 15 | <0.0001 | ||
| 3/4 | 1.4 | 1.9 | −3.2 | 6.0 | 0.75 | ||||||
| CT%ca | 887 | 2 | 34 | 39 | 12 | 4/2 | 8.0 | 2.0 | 3.3 | 13 | 0.0002 |
| 3 and 4 | 89 | 44 | 6.9 | 3/2 | 6.9 | 2.1 | 2.0 | 12 | 0.0031 | ||
| 4/3 | 1.1 | 1.9 | −3.3 | 5.6 | 0.82 | ||||||
| CC%ct | 983 | 2 | 34 | 61 | 18 | 2/3 | 16 | 3.0 | 8.6 | 23 | <0.0001 |
| 3 and 4 | 89 | 46 | 11 | 2/4 | 15 | 2.9 | 8.1 | 22 | <0.0001 | ||
| 4/3 | 0.8 | 2.8 | −5.7 | 7.4 | 0.95 | ||||||
| CC%ca | 997 | 2 | 34 | 53 | 18 | 2/4 | 14 | 3.0 | 6.9 | 21 | <0.0001 |
| 3 and 4 | 89 | 42 | 12 | 2/3 | 8.6 | 3.2 | 1.1 | 16 | 0.0211 | ||
| 3/4 | 5.5 | 2.9 | −1.3 | 12 | 0.14 | ||||||
| TT + CT%ct | 983 | 2 | 34 | 39 | 18 | 3/2 | 16 | 3.0 | 8.6 | 23 | <0.0001 |
| 3 and 4 | 89 | 54 | 11 | 4/2 | 15 | 2.9 | 8.1 | 22 | <0.0001 | ||
| 3/4 | 0.8 | 2.8 | −5.7 | 7.4 | 0.9499 | ||||||
| TT + CT%ca | 997 | 2 | 34 | 47 | 18 | 4/2 | 14 | 3.0 | 6.9 | 21 | <0.0001 |
| 89 | 58 | 12 | 3/2 | 8.6 | 3.2 | 1.1 | 16 | 0.0211 | |||
| 4/3 | 5.5 | 2.9 | −1.3 | 12 | 0.1410 | ||||||
| RRTT | 373 | 2 | 29 | 1.8 | 1.7 | 2/3 | 0.6 | 0.3 | −0.1 | 1.2 | 0.11 |
| 3 and 4 | 89 | 1.4 | 0.9 | 2/4 | 0.3 | 0.3 | −0.3 | 1.0 | 0.42 | ||
| 4/3 | 0.2 | 0.3 | −0.4 | 0.8 | 0.60 | ||||||
| RRCT | 137 | 2 | 34 | 1.3 | 0.8 | 2/3 | 0.3 | 0.1 | 0.1 | 0.6 | 0.0031 |
| 3 and 4 | 89 | 1.1 | 0.2 | 2/4 | 0.3 | 0.1 | 0.03 | 0.5 | 0.0244 | ||
| 4/3 | 0.1 | 0.1 | −0.1 | 0.3 | 0.65 | ||||||
| RRTT + CT | 156 | 2 | 34 | 1.4 | 0.8 | 2/3 | 0.4 | 0.1 | 0.2 | 0.6 | 0.0007 |
| 3 and 4 | 89 | 1.1 | 0.2 | 2/4 | 0.3 | 0.1 | 0.04 | 0.5 | 0.0200 | ||
| 4/3 | 0.1 | 0.1 | −0.1 | 0.4 | 0.40 | ||||||
| RRCC | −84 | 2 and 3 | 74 | 0.9 | 0.2 | 3/4 | 0.2 | 0.04 | 0.1 | 0.2 | 0.0001 |
| 4 | 49 | 0.9 | 0.2 | 3/2 | 0.1 | 0.04 | 0.03 | 0.2 | 0.0041 | ||
| 2/4 | 0.02 | 0.04 | −0.1 | 0.1 | 0.81 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.-F.; Young, L.; Yu, C.H.; Shiao, S.P.K. A Meta-Prediction of Methylenetetrahydrofolate-Reductase Polymorphisms and Air Pollution Increased the Risk of Ischemic Heart Diseases Worldwide. Int. J. Environ. Res. Public Health 2018, 15, 1453. https://doi.org/10.3390/ijerph15071453
Chen Z-F, Young L, Yu CH, Shiao SPK. A Meta-Prediction of Methylenetetrahydrofolate-Reductase Polymorphisms and Air Pollution Increased the Risk of Ischemic Heart Diseases Worldwide. International Journal of Environmental Research and Public Health. 2018; 15(7):1453. https://doi.org/10.3390/ijerph15071453
Chicago/Turabian StyleChen, Zhao-Feng, Lufei Young, Chong Ho Yu, and S. Pamela K. Shiao. 2018. "A Meta-Prediction of Methylenetetrahydrofolate-Reductase Polymorphisms and Air Pollution Increased the Risk of Ischemic Heart Diseases Worldwide" International Journal of Environmental Research and Public Health 15, no. 7: 1453. https://doi.org/10.3390/ijerph15071453