
Adaptability and Stability Analysis of Commercial Cultivars, Experimental Hybrids and Lines under Natural Fall Armyworm Infestation in Zimbabwe Using Different Stability Models

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm for Testing
2.2. Trial Sites, Experimental Design and Agronomic Management
2.3. Data Collection and Analysis
3. Results
3.1. Analysis of Genotype by Environment Effects for Grain Yield, Fall Armyworm Resistance and Related Traits of Hybrids/OPVs Using the AMMI Model
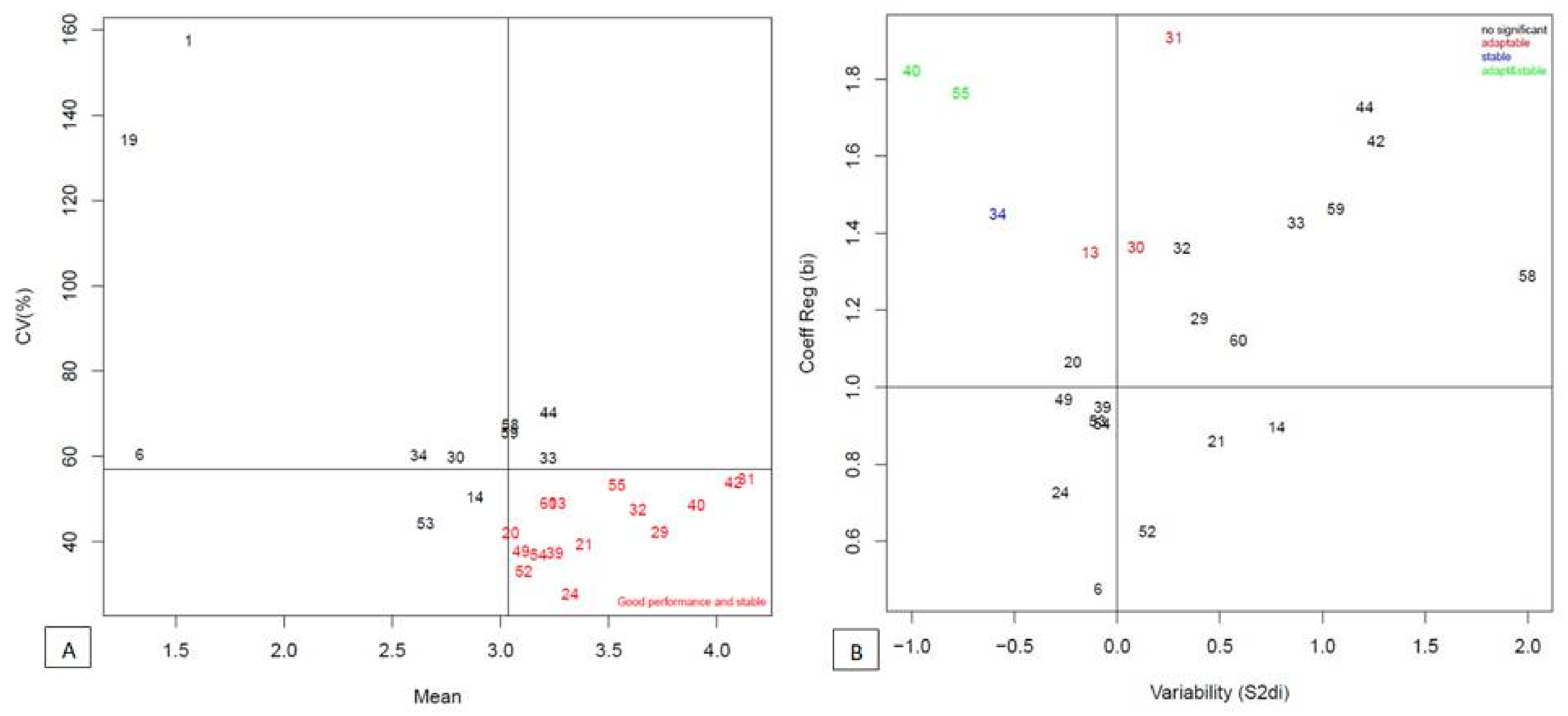
3.2. Mean Grain Yield Performance of Hybrids/OPVs and Stability Analysis
3.3. Identification of the Best Screening Environments for Grain Yield Potential under Natural Fall Armyworm Infestation in Zimbabwe
3.4. Inbred Lines Exhibiting High Adaptation and Foliar Fall Armyworm Resistance Stability across Environments with Natural Fall Armyworm Infestation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Day, R.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa; Evidence Note (2); CABI: Oxfordshire, UK, 2017. [Google Scholar]
- Kansiime, M.K.; Mugambi, I.; Rwomushana, I.; Nunda, W.; Lamontagne-Godwin, J.; Rware, H.; Phiri, N.A.; Chipabika, G.; Ndlovu, M.; Day, R. Farmer perception of fall armyworm (Spodoptera frugiderda J.E. Smith) and farm-level management practices in Zambia. Pest. Manag. Sci. 2019, 75, 2840–2850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matova, P.M.; Kamutando, C.N.; Magorokosho, C.; Kutywayo, D.; Gutsa, F.; Labuschagne, M. Fall-armyworm invasion, control practices and prospects of breeding fall-armyworm-resistant maize in Sub-Saharan Africa: A review. Crop Sci. 2020, 60, 2951–2970. [Google Scholar] [CrossRef] [PubMed]
- Kasoma, C.; Shimelis, H.; Laing, M.; Shayanowako, A.I.T.; Mathew, I. Screening of inbred lines of tropical maize for resistance to fall armyworm, and for yield and yield-related traits. Crop Prot. 2020, 136, 105218. [Google Scholar] [CrossRef]
- CIMMYT. Highlighted impacts from the CGIAR Research Program on MAIZE. In Maize Annual Report 2020; CIMMYT: Texcoco, Mexico, 2020. [Google Scholar]
- De Oliveira, E.J.; de Freitas, J.P.X.; de Jesus, O.N. AMMI analysis of the adaptability and yield stability of yellow passion fruit varieties. Sci. Agric. 2014, 71, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Wilsie, C.P. Crop Adaptation and Distribution; W.H. Freeman and Company: San Franscisco, CA, USA; London, UK, 1962. [Google Scholar]
- Becker, H.C.; Léon, J. Stability analysis in plant breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Pacheco, Á.; Vargas, M.; Alvarado, G.; Rodríguez, F.; Crossa, J.; Burgueño, J. GEA-R (Genotype × Environment Analysis with R for Windows) Version 4.1, 2015. CIMMYT Research Data & Software Repository Network, V16. Available online: https://hdl.handle.net/11529/10203 (accessed on 17 April 2019).
- Matova, P.M.; Kamutando, C.N.; Kutywayo, D.; Magorokosho, C.; Labuschagne, M. Fall armyworm response of maize parental lines, experimental hybrids, and commercial cultivars in southern Africa. Agronomy 2022, 12, 1463. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; Feed the Future: Mexico City, Mexico, 2018. [Google Scholar]
- Magorokosho, C.; Vivek, B.J.; MacRobert, J.F. Characterization of Maize Germplasm Grown in Eastern and Southern Africa: Results of the 2007 Regional Trials Coordinated by CIMMYT; CIMMYT: Harare, Zimbabwe, 1998. [Google Scholar]
- Masuka, B.P.; van Biljon, A.; Cairns, J.E.; Das, B.; Labuschagne, M.; MacRobert, J.; Makumbi, D.; Magorokosho, C.; Zaman-Allah, M.; Ogugo, V.; et al. Genetic diversity among selected elite CIMMYT maize hybrids in east and Southern Africa. Crop Sci. 2017, 57, 2395–2404. [Google Scholar] [CrossRef] [Green Version]
- VSN International. Genstats for Windows, 18th ed.; VSN International: Hemel Hempstead, UK, 2017. [Google Scholar]
- Alvarado, G.; Rodríguez, F.M.; Pacheco, A.; Burgueño, J.; Crossa, J.; Vargas, M.; Pérez-Rodrígues, P.; Lopez-Cruz, M.A. META-R: A software to analyze data from multi-environment plant breeding trials. Crop J. 2020, 8, 745–756. [Google Scholar] [CrossRef]
- Gauch, H.G. Statistical analysis of yield trials by AMMI and GGE. Crop Sci. 2006, 46, 1488–1500. [Google Scholar] [CrossRef]
- Gauch, H.G.; Zobel, R.W. Optimal replication in selection experiments. Crop Sci. 1996, 36, 838–843. [Google Scholar] [CrossRef]
- Purchase, J.L.; Hatting, H.; van Deventer, C.S. Genotype × environment interaction of winter wheat (Triticum aestivum L.) in South Africa: II. Stability analysis of yield performance. South Afr. J. Plant Soil 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Binns, M.R. A superiority measure of cultivar performance for cultivar × location data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Matova, P.M.; Gasura, E. Yield and stability of new cowpea varieties in Zimbabwe. Afr. Crop Sci. J. 2018, 26, 277. [Google Scholar] [CrossRef] [Green Version]
- Gasura, E.; Setimela, P.S.; Souta, C.M. Evaluation of the performance of sorghum genotypes using GGE biplot. Can. J. Plant Sci. 2015, 95, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, E.J.; de Godoy, I.J. Pod yield stability analysis of runner peanut lines using AMMI. Crop Breed. Appl. Biotechnol. 2006, 6, 310–317. [Google Scholar] [CrossRef]
- Weedon, O.D.; Finckh, M.R. Heterogeneous winter wheat populations differ in yield stability depeneding on their genetic background and management system. Sustainability 2019, 11, 6172. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype Code | Name | Source | Year of Release | Production Region | Line Code | Line Name | Source | Type Status/Origin |
|---|---|---|---|---|---|---|---|---|
| G1 | Salisbury white | CBI | Not Clear | Zimbabwe and ESA | G25 | CLHP0005 | HarvestPlus | local |
| G6 | R215 | CBI | 1974 | Zimbabwe | G27 | CML304 | CIMMYT | local |
| G13 | ZS265 | CBI | 2011 | Zimbabwe | G42 | CZL1227 | CIMMYT | local |
| G14 | ZS269 | CBI | 2014 | Zimbabwe | G28 | CML444 | CIMMYT | local |
| G19 | SR52 | CBI | 1962 | Zimbabwe and ESA | G57 | CML543 | CIMMYT | local |
| G20 | ZS242A | CBI | 2015 | Zimbabwe and ESA | G34 | CML395 | CIMMYT | local |
| G21 | ZS246A | CBI | 2016 | Zimbabwe and ESA | G48 | CML334 | CIMMYT | local |
| G24 | 113WH330 | CBI | Experimental | Zimbabwe | G33 | CML312 | CIMMYT | local |
| G29 | CZH1258 | CIMMYT | Experimental | N/A | G36 | CLHP00478 | HarvestPlus | local |
| G30 | NTS51 | NTS | 2014 | Zimbabwe | G51 | CML139 | CIMMYT | local |
| G31 | PAN53 | PANNAR | 2007 | Zimbabwe and ESA | G52 | CML571 | CIMMYT | local |
| G32 | PAN4M-23 | PANNAR | - | Zimbabwe and ESA | G24 | CLHP0003 | HarvestPlus | local |
| G33 | PAN-7M-81 | PANNAR | 2013 | Zimbabwe and ESA | G60 | CimExp54 | CIMMYT | local |
| G34 | PHB30G19 | PIONEER | 2008 | Zimbabwe and ESA | G50 | CML67 | CIMMYT | Exotic |
| G39 | Manjanja MN421 | Mukushi | 2015 | Zimbabwe, South Africa, Zambia | G55 | CML121 | CIMMYT | Exotic |
| G40 | Mutsa MN521 | Mukushi | 2014 | Zimbabwe, South Africa, Zambia | G47 | CML338 | CIMMYT | Exotic |
| G42 | Mukwa | Mukushi | 2016 | Zimbabwe, South Africa, Zambia | G53 | CML346 | CIMMYT | Exotic |
| G44 | ZAP61 | Agriseeds | 2008 | Zimbabwe and ESA | G9 | SV1P | CBI-DR & SS | local |
| G49 | CML338/CML334 | CIMMYT | Experimental | N/A | G49 | CML331 | CIMMYT | Exotic |
| G52 | CIM52/CML139 | CIMMYT | Experimental | N/A | G30 | CML491 | CIMMYT | local |
| G53 | CIM53/CML345 | CIMMYT | Experimental | N/A | G61 | CZL1112 | CIMMYT | local |
| G54 | CIM54/CML334 | CIMMYT | Experimental | N/A | G38 | DPTY…*9 | HarvestPlus | local |
| G55 | CIM55/CML334 | CIMMYT | Experimental | N/A | G46 | CML539 | CIMMYT | local |
| G58 | CIM58/CML121 | CIMMYT | Experimental | N/A | G40 | CZL1315 | CIMMYT | local |
| G59 | CML543/CML334 | CIMMYT | Experimental | N/A | G23 | WW01408 | CBI-DR & SS | local |
| G60 | CML571/CML338 | CIMMYT | Experimental | N/A | G18 | HX482P | CBI-DR & SS | local |
| Precipitation | |||||||
|---|---|---|---|---|---|---|---|
| Location | Management | Altitude | Latitude | Longitude | 2018/2019 | 2019/2020 | Fertilisers (NPK) ha−1 |
| m | -mm- | ||||||
| CIM-Harare | NI | 1506 | 17°48′ S | 31°85′ E | 557.2 | 547.3 | 166:24.5:23.2 |
| RARS | NI | 1341 | 17°14′ S | 31°14′ E | 631.8 | 543.8 | 166:24.5:23.2 |
| Chisumbanje | NI | 421 | 20°05′ S | 32°15′ E | 441.9 | 434.8 | 166:24.5:23.2 |
| Chiredzi | NI | 409 | 21°01′ S | 21°25′ E | 416.5 | 419.2 | 166:24.5:23.2 |
| DR&SS-Harare | NI + Managed | 1506 | 17°48′ S | 31°03′ E | 502.7 | 436.3 | 166:24.5:23.2 |
| Gwebi VTC | NI | 1448 | 17°41′ S | 30°32′ E | 571.5 | 542.5 | 166:24.5:23.2 |
| KD-CRI | NI | 1149 | 18°94′ S | 29°25′ E | 555.6 | 474.8 | 28:24.5:23.2 |
| (A) Hybrids/OPVs | GYD | FFAWD | EFAWD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | DF | SS | MS | DF | SS | MS | DF | SS | MS |
| Treatments | 233 | 1405.5 | 6.03 *** | 285 | 49,044 | 172.10 *** | 155 | 565.6 | 3.65 *** |
| Genotypes | 25 | 255.5 | 10.22 *** | 25 | 8283 | 331.30 *** | 25 | 91.5 | 3.66 *** |
| Environments | 8 | 519.6 | 64.95 *** | 10 | 25,920 | 2592.00 *** | 5 | 238.8 | 47.75 *** |
| Block | 9 | 18 | 2.00 | 11 | 355 | 32.3 | 6 | 9.6 | 1.60 |
| Interactions | 196 | 630.4 | 3.22 *** | 250 | 14841 | 59.40 ** | 123 | 235.4 | 1.91 *** |
| IPCA 1 | 32 | 283.7 | 8.86 *** | 34 | 4604 | 135.40 ** | 29 | 97.2 | 3.35 *** |
| IPCA 2 | 30 | 117.8 | 3.93 *** | 32 | 2901 | 90.60 *** | 27 | 61.8 | 2.29 ** |
| Residuals | 134 | 229 | 1.71 | 184 | 7337 | 39.9 | 67 | 76.3 | 1.14 |
| Error | 199 | 342.6 | 1.72 | 275 | 11,339 | 41.2 | 147 | 150.9 | 1.03 |
| (B) Inbred lines | |||||||||
| Treatments | 181 | 99.47 | 0.55 *** | 259 | 1206.5 | 4.66 *** | 129 | 375.3 | 2.91 ** |
| Genotypes | 25 | 39.31 | 1.52 *** | 25 | 524.2 | 20.97 *** | 25 | 137 | 5.48 *** |
| Environments | 6 | 23.2 | 3.87 *** | 9 | 427.8 | 47.54 *** | 4 | 60.4 | 15.11 *** |
| Block | 7 | 3.08 | 0.44 | 10 | 18.4 | 1.84 * | 5 | 0.9 | 0.19 |
| Interactions | 136 | 36.96 | 0.27 | 224 | 254.4 | 1.36 * | 94 | 177.8 | 1.89 |
| IPCA 1 | 30 | 14.71 | 0.49 * | 33 | 84.7 | 2.57 *** | 28 | 104.2 | 3.72 ** |
| IPCA 2 | 28 | 9.43 | 0.37 | 31 | 42.6 | 1.37 * | 26 | 40 | 1.54 |
| Residuals | 78 | 12.81 | 0.16 | 160 | 127.1 | 0.8 | 40 | 33.6 | 0.84 |
| Error | 124 | 37.43 | 0.3 | 233 | 194.6 | 0.84 | 102 | 186.8 | 1.83 |
| AMMI Stability Model | Mean Square Deviation Model | Lin and Binns Model | Eberhart and Russel Model | Wricke’s Ecovalence Model | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype Number | Genotype Code | Name | IPCAg1 | IPCAg2 | Mean | GYD Rank | ASV | ASV Rank | YSI | YSI Rank | Sd | Sd Rank | Cultivar Superiority | Cs Rank | Static Stability | Ss Rank | Wricke’s Ecovalence | We Rank |
| 1 | G1 | Salisbury White | −2.10 | 0.32 | 1.68 | 24 | 3.27 | 26 | 50 | 26 | 2.45 | 26 | 10.79 | 24 | * | * | 53.19 | 26 |
| 2 | G6 | R215 | −0.07 | −0.20 | 1.30 | 25 | 0.23 | 1 | 26 | 13 | 0.81 | 1 | 11.43 | 25 | 0.65 | 1 | 6.63 | 10 |
| 3 | G13 | ZS265 | −0.45 | −0.46 | 3.29 | 9 | 0.84 | 12 | 21 | 9 | 1.60 | 14 | 3.42 | 5 | 2.55 | 14 | 6.73 | 11 |
| 4 | G14 | ZS269 | −0.58 | −0.63 | 2.88 | 20 | 1.10 | 17 | 37 | 22 | 1.45 | 10 | 5.14 | 17 | 2.11 | 10 | 11.64 | 18 |
| 5 | G19 | SR52 | −0.24 | 1.48 | 0.95 | 26 | 1.53 | 23 | 49 | 25 | 1.72 | 16 | 13.13 | 26 | * | * | 19.95 | 23 |
| 6 | G20 | ZS242A | −0.43 | −0.56 | 3.05 | 17 | 0.86 | 13 | 30 | 18 | 1.28 | 8 | 4.21 | 12 | 1.65 | 8 | 4.90 | 3 |
| 7 | G21 | ZS246A | −0.80 | −0.36 | 3.39 | 7 | 1.30 | 20 | 27 | 15 | 1.33 | 9 | 3.51 | 7 | 1.78 | 9 | 10.56 | 16 |
| 8 | G24 | 113WH330 | 0.20 | 0.27 | 3.30 | 8 | 0.41 | 3 | 11 | 1 | 0.93 | 2 | 4.49 | 14 | 0.86 | 2 | 5.14 | 4 |
| 9 | G29 | CZH128 | 0.75 | 0.04 | 3.72 | 4 | 1.17 | 19 | 23 | 10 | 1.58 | 11 | 3.90 | 9 | 2.49 | 11 | 9.66 | 14 |
| 10 | G30 | NTS51 | 0.35 | −0.17 | 2.77 | 21 | 0.57 | 6 | 27 | 16 | 1.67 | 15 | 5.50 | 19 | 2.80 | 15 | 7.69 | 12 |
| 11 | G31 | PAN53 | 1.03 | −0.22 | 4.14 | 1 | 1.62 | 24 | 25 | 12 | 2.26 | 24 | 3.12 | 4 | 5.12 | 23 | 16.75 | 21 |
| 12 | G32 | PAN4M-23 | −0.23 | −0.96 | 3.64 | 5 | 1.02 | 15 | 20 | 6 | 1.73 | 17 | 3.01 | 3 | 2.98 | 16 | 10.13 | 15 |
| 13 | G33 | PAN7M-81 | 0.42 | 0.49 | 3.23 | 10 | 0.82 | 10 | 20 | 7 | 1.92 | 20 | 4.93 | 16 | 3.69 | 19 | 13.93 | 19 |
| 14 | G34 | PHB30G19 | 0.44 | −0.09 | 2.60 | 23 | 0.68 | 7 | 30 | 19 | 1.58 | 12 | 5.87 | 22 | 2.50 | 12 | 3.53 | 1 |
| 15 | G39 | Manjanja-MN421 | 0.07 | 0.44 | 3.22 | 13 | 0.45 | 4 | 17 | 3 | 1.22 | 7 | 4.20 | 11 | 1.48 | 7 | 6.07 | 6 |
| 16 | G40 | Mutsa-MN521 | 0.44 | −0.01 | 3.91 | 3 | 0.68 | 8 | 11 | 2 | 1.90 | 19 | 2.41 | 1 | 3.61 | 18 | 6.32 | 7 |
| 17 | G42 | Mukwa | 0.56 | 0.75 | 4.08 | 2 | 1.15 | 18 | 20 | 8 | 2.20 | 23 | 2.95 | 2 | 4.82 | 22 | 19.98 | 24 |
| 18 | G44 | ZAP61 | 1.01 | −0.44 | 3.22 | 11 | 1.63 | 25 | 36 | 21 | 2.27 | 25 | 5.43 | 18 | 5.14 | 24 | 19.16 | 22 |
| 19 | G49 | CML338/CML334 | −0.17 | 0.31 | 3.10 | 16 | 0.41 | 2 | 18 | 5 | 1.17 | 4 | 4.43 | 13 | 1.37 | 4 | 4.77 | 2 |
| 20 | G52 | CimExp52/CML139 | −0.66 | 0.18 | 3.11 | 15 | 1.03 | 16 | 31 | 20 | 1.03 | 3 | 4.53 | 15 | 1.06 | 3 | 9.12 | 13 |
| 21 | G53 | CimExp53/CML345 | −0.31 | 0.08 | 2.64 | 22 | 0.49 | 5 | 27 | 17 | 1.18 | 5 | 5.71 | 21 | 1.38 | 5 | 5.39 | 5 |
| 22 | G54 | CimExp54/CML334 | −0.48 | 0.03 | 3.14 | 14 | 0.75 | 9 | 23 | 11 | 1.18 | 6 | 3.87 | 8 | 1.39 | 6 | 6.42 | 8 |
| 23 | G55 | CimExp55/CML334 | 0.53 | 0.02 | 3.54 | 6 | 0.83 | 11 | 17 | 4 | 1.89 | 18 | 3.43 | 6 | 3.56 | 17 | 6.57 | 9 |
| 24 | G58 | CimExp58/CMC121 | 0.50 | −1.16 | 3.02 | 18 | 1.40 | 21 | 39 | 23 | 2.05 | 22 | 6.05 | 23 | 4.22 | 21 | 20.66 | 25 |
| 25 | G59 | CML543/CML334 | 0.80 | 0.73 | 3.02 | 19 | 1.43 | 22 | 41 | 24 | 2.00 | 21 | 5.67 | 20 | 3.98 | 20 | 15.37 | 20 |
| 26 | G60 | CML571/CML338 | −0.58 | 0.10 | 3.22 | 12 | 0.91 | 14 | 26 | 14 | 1.58 | 13 | 3.92 | 10 | 2.50 | 13 | 11.03 | 17 |
| Genotype Number | Genotype Code | Name | Grain Yield | Grain Yield Rank |
|---|---|---|---|---|
| 1 | G9 | SV1P | 1.37 | 1 |
| 2 | G18 | HX482P | 0.10 | 25 |
| 3 | G23 | WW01408 | 0.07 | 26 |
| 4 | G24 | CLHP0003 | 0.52 | 15 |
| 5 | G25 | CLHP0005 | 0.60 | 12 |
| 6 | G27 | CML304 | 0.79 | 5 |
| 7 | G28 | CML444 | 0.31 | 21 |
| 8 | G30 | CML491 | 1.30 | 2 |
| 9 | G33 | CML312 | 0.45 | 18 |
| 10 | G34 | CML395 | 0.37 | 19 |
| 11 | G36 | CLHP00478 | 0.31 | 20 |
| 12 | G38 | DPTY…*9 | 1.15 | 3 |
| 13 | G40 | CZL1315 | 0.71 | 8 |
| 14 | G42 | CZL1227 | 0.67 | 10 |
| 15 | G46 | CML539 | 0.49 | 17 |
| 16 | G47 | CML338 | 0.78 | 6 |
| 17 | G48 | CML334 | 0.71 | 7 |
| 18 | G49 | CML331 | 0.59 | 13 |
| 19 | G50 | CML67 | 0.63 | 11 |
| 20 | G51 | CML139 | 0.30 | 22 |
| 21 | G52 | CML571 | 0.26 | 23 |
| 22 | G53 | CML346 | 0.50 | 16 |
| 23 | G55 | CML121 | 0.88 | 4 |
| 24 | G57 | CML543 | 0.13 | 24 |
| 25 | G60 | Cim60Exp | 0.69 | 9 |
| 26 | G61 | CZL1112 | 0.54 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matova, P.M.; Kamutando, C.N.; Mutari, B.; Magorokosho, C.; Labuschagne, M. Adaptability and Stability Analysis of Commercial Cultivars, Experimental Hybrids and Lines under Natural Fall Armyworm Infestation in Zimbabwe Using Different Stability Models. Agronomy 2022, 12, 1724. https://doi.org/10.3390/agronomy12071724
Matova PM, Kamutando CN, Mutari B, Magorokosho C, Labuschagne M. Adaptability and Stability Analysis of Commercial Cultivars, Experimental Hybrids and Lines under Natural Fall Armyworm Infestation in Zimbabwe Using Different Stability Models. Agronomy. 2022; 12(7):1724. https://doi.org/10.3390/agronomy12071724
Chicago/Turabian StyleMatova, Prince M., Casper N. Kamutando, Bruce Mutari, Cosmos Magorokosho, and Maryke Labuschagne. 2022. "Adaptability and Stability Analysis of Commercial Cultivars, Experimental Hybrids and Lines under Natural Fall Armyworm Infestation in Zimbabwe Using Different Stability Models" Agronomy 12, no. 7: 1724. https://doi.org/10.3390/agronomy12071724