
In Silico Identification of lncRNAs Regulating Sperm Motility in the Turkey (Meleagris gallopavo L.)

 ,
,  , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Sequencing, Mapping and Expression
2.2. lncRNA Identification
2.3. Differentially Expressed lncRNAs


2.4. TRANS-Acting Genes
2.5. Direct Interactions
2.6. Real-Time PCR-Validation of the Expression of Selected lncRNAs
3. Discussion
3.1. The Regulations between Testes and Post-Testicular Areas of Spermatogenesis
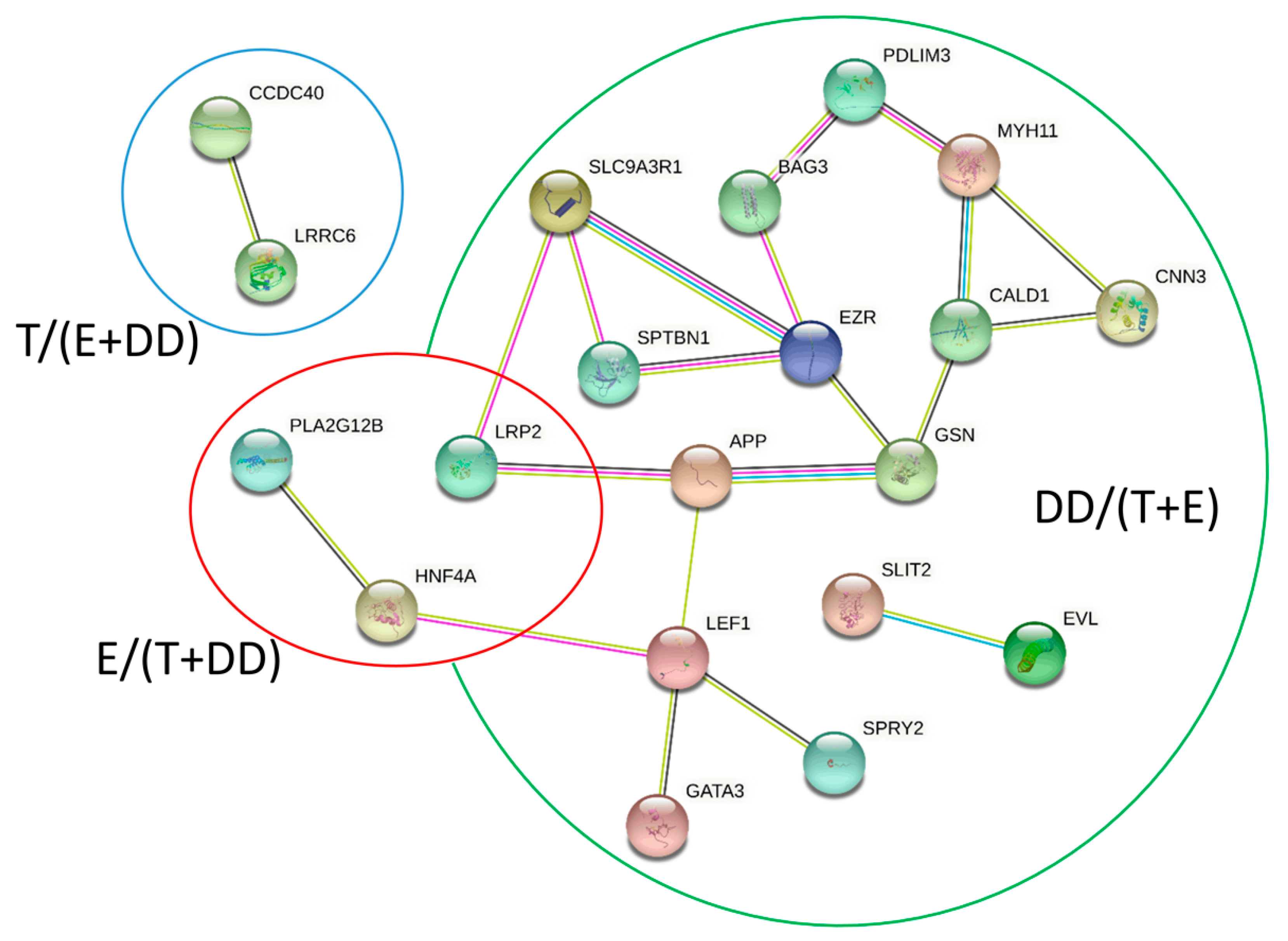
Testes (Group T/(E + DD))
3.2. Post-Testicular Development
3.2.1. Epididymis (Group E/(T + DD))
3.2.2. Ductus Deferens (Group DD/(T + E))
4. Materials and Methods
4.1. Tissue Collection
4.2. RNA Isolation, Library Preparation and Sequencing
4.3. In Silico Preprocessing, Mapping and Expression Analyses
4.4. Expression Profiling and Differentially Expressed Genes
4.5. The lncRNA Identification
4.6. The lncRNA Interactions with Sperm Motility-Related DEGs
4.7. Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACVR1 | Activin receptor type-1 |
| ANXA1 | Annexin A1 |
| APP | Amyloid-beta precursor protein |
| ARMC4 | Armadillo repeat containing 4 |
| ARPC1-5 | Actin related protein 2/3 complex subunit from 1 to 5 |
| ATP1A1 | ATPase Na+/K+ transporting subunit alpha 1 |
| BAG3 | BCL2 associated athanogene 3 |
| CCDC40 | Coiled-coil domain-containing protein 40 |
| CDH20 | Cadherin 20 |
| CDHR2 | Cadherin related family member 2 |
| CFAP221 | Cilia and flagella-associated protein 221 |
| CFAP69 | Cilia and flagella associated protein 69 |
| CGNL1 | Cingulin Like 1 |
| CLGN | Calmegin |
| CNN3 | Calponin 3 |
| CTBP1 | C-terminal binding protein 1 |
| DEUP1 | Deuterosome assembly protein 1 |
| DNAH5 | Dynein axonemal heavy chain 5 |
| DNAH7 | Dynein axonemal heavy chain 7 |
| EVL | Ena/vasodilator-stimulated phosphoprotein-like protein |
| EZR | Ezrin |
| FAT1 | FAT atypical cadherin 1 |
| GATA3 | GATA binding protein 3 |
| GSN | Gelsolin |
| HEXB | Hexosaminidase subunit beta |
| HNF4A | Hepatocyte nuclear factor 4 alpha |
| HPGDS | Hematopoietic prostaglandin D synthase |
| ITGB1 | Integrin subunit beta 1 |
| KRT19 | Keratin 19 |
| LEF1 | Lymphoid enhancer binding factor 1 |
| LRP2 | LDL receptor related protein 2 |
| LRRC6 | Leucine rich repeat containing 6 |
| MET | MET proto-oncogene. receptor tyrosine kinase |
| MICAL2 | Microtubule associated monooxygenase. calponin and LIM domain containing 2 |
| MYH11 | Myosin Heavy Chain 11 |
| NDC80 | NDC80. kinetochore complex component |
| NEDD9 | Neural precursor cell expressed, developmentally down-regulated 9 |
| NET1 | Neuroepithelial transforming gene 1 |
| NRP1 | Neuropilin 1 |
| PACSIN2 | Protein kinase C and casein kinase substrate in neurons 2 |
| PDLIM3 | PDZ and LIM domain 3 |
| PIH1D3 | PIH1 domain containing 3 |
| PLA2G12B | Phospholipase A2 group XIIB |
| PLK4 | Polo like kinase 4 |
| POF1B | POF1B. actin binding protein |
| RDH10 | Retinol dehydrogenase 10 |
| ROPN1L | Rhophilin associated tail protein 1 like |
| RSPH9 | Radial spoke head 9 homolog |
| SDCBP | Syndecan binding protein |
| SLC9A3R1 | SLC9A3 regulator 1 |
| SLIT2 | Slit guidance ligand 2 |
| SORBS1 | Sorbin and SH3 domain containing 1 |
| SPRY2 | Sprouty RTK signaling antagonist 2 |
| SPTBN1 | Spectrin beta, non-erythrocytic 1 |
| TTC26 | Tetratricopeptide repeat domain 26 |
| TWF1 | Twinfilin actin binding protein 1 |
| VEGFA | Vascular endothelial growth factor A |
| ZMYND10 | Zinc finger MYND-type containing 10 |
References
- Pardyak, L.; Kaminska, A.; Brzoskwinia, M.; Hejmej, A.; Kotula-Balak, M.; Jankowski, J.; Ciereszko, A.; Bilinska, B. Differences in aromatase expression and steroid hormone concentrations in the reproductive tissues of male domestic turkeys (Meleagris gallopavo) with white and yellow semen. Br. Poult. Sci. 2018, 59, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Pardyak, L.; Kaminska, A.; Brzoskwinia, M.; Hejmej, A.; Kotula-Balak, M.; Jankowski, J.; Ciereszko, A.; Bilinska, B. Differential expression of cell-cell junction proteins in the testis, epididymis, and ductus deferens of domestic turkeys (Meleagris gallopavo) with white and yellow semen. Poult. Sci. 2020, 99, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Hess, R.A.; Sharpe, R.M.; Hinton, B.T. Estrogens and development of the rete testis, efferent ductules, epididymis and vas deferens. Differentiation 2021, 118, 41–71. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Brym, P.; Pareek, C.S.; Mogielnicka-Brzozowska, M.; Paukszto, Ł.; Jastrzębski, J.P.; Wasilewska-Sakowska, K.; Mańkowska, A.; Sobiech, P.; Żukowski, K. Transcriptome analysis of boar spermatozoa with different freezability using RNA-Seq. Theriogenology 2020, 142, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Słowińska, M.; Paukszto, Ł.; Jastrzębski, J.P.; Bukowska, J.; Kozłowski, K.; Jankowski, J.; Ciereszko, A. Transcriptome analysis of turkey (Meleagris gallopavo) reproductive tract revealed key pathways regulating spermatogenesis and post-testicular sperm maturation. Poult. Sci. 2020, 99, 6094–6118. [Google Scholar] [CrossRef]
- Fraser, L.; Paukszto, Ł.; Mańkowska, A.; Brym, P.; Gilun, P.; Jastrzębski, J.P.; Pareek, C.S.; Kumar, D.; Pierzchała, M. Regulatory potential of long non-coding rnas (LncRNAs) in boar spermatozoa with good and poor freezability. Life 2020, 10, 300. [Google Scholar] [CrossRef]
- Joshi, M.; Rajender, S. Long non-coding RNAs (lncRNAs) in spermatogenesis and male infertility. Reprod. Biol. Endocrinol. 2020, 18, 103. [Google Scholar] [CrossRef]
- Harvey, S.; Scanes, C.G.; Phillips, J.G. Advances in Experimental Medicine and Biology. In Avian Reproduction; Sasanami, T., Ed.; Springer: Singapore, 2017; Volume 1001, ISBN 978-981-10-3974-4. [Google Scholar]
- Johnson, L.; Varner, D.D.; Roberts, M.E.; Smith, T.L.; Keillor, G.E.; Scrutchfield, W.L. Efficiency of spermatogenesis: A comparative approach. Anim. Reprod. Sci. 2000, 60–61, 471–480. [Google Scholar] [CrossRef]
- Oliva, R.; Mezquita, C. Marked differences in the ability of distinct protamines to disassemble nucleosomal core particles in vitro. Biochemistry 1986, 25, 6508–6511. [Google Scholar] [CrossRef]
- Sprando, R.L.; Russell, L.D. Spermiogenesis in the bluegill (Lepomis macrochirus): A study of cytoplasmic events including cell volume changes and cytoplasmic elimination. J. Morphol. 1988, 198, 165–177. [Google Scholar] [CrossRef]
- Aire, T.A.; Olowo-okorun, M.O.; Ayeni, J.S. The seminiferous epithelium in the guinea fowl (Numida meleagris). Cell Tissue Res. 1980, 205, 319–325. [Google Scholar] [CrossRef] [PubMed]
- De Reviers, M.; Richetin, C.; Brillard, J.P. LE DÉVELOPPEMENT TESTICULAIRE CHEZ LE COQ. II.—MORPHOLOGIE DE L’ÉPITHÉLIUM SÉMINIFÈRE ET ÉTABLISSEMENT DE LA SPERMATOGENÈSE. Ann. Biol. Anim. Biochim. Biophys. 1971, 11, 531–546. [Google Scholar] [CrossRef]
- Gunawardana, V.E. Stages of spermatids in the domestic fowl: A light microscope study using Araldite sections. J. Anat. 1977, 123, 351–360. [Google Scholar] [PubMed]
- Tiba, T.; Yoshida, K.; Miyake, M.; Tsuchiya, K.; Kita, I.; Tsubota, T. Regularities and Irregularities in the Structure of the Seminiferous Epithelium in the Domestic Fowl (Gallus domesticus). Anat. Histol. Embryol. 1993, 22, 241–253. [Google Scholar] [CrossRef]
- Lin, M.; Jones, R.C.; Blackshaw, A.W. The cycle of the seminiferous epithelium in the Japanese quail (Coturnix coturnix japonica) and estimation of its duration. Reproduction 1990, 88, 481–490. [Google Scholar] [CrossRef]
- Lin, M.; Jones, R.C. Spermiogenesis and spermiation in the Japanese quail (Coturnix coturnix japonica). J. Anat. 1993, 183 Pt 3, 525–535. [Google Scholar]
- Freitas, M.J.; Vijayaraghavan, S.; Fardilha, M. Signaling mechanisms in mammalian sperm motility. Biol. Reprod. 2017, 96, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Gunes, S.; Sengupta, P.; Henkel, R.; Alguraigari, A.; Sinigaglia, M.M.; Kayal, M.; Joumah, A.; Agarwal, A. Microtubular dysfunction and male infertility. World J. Mens. Health 2020, 38, 9–23. [Google Scholar] [CrossRef]
- Martins, M.C.; Gonçalves, L.M.; Nonato, A.; Travençolo, B.A.N.; Alves, B.G.; Beletti, M.E. Sperm head morphometry and chromatin condensation are in constant change at seminiferous tubules, epididymis, and ductus deferens in bulls. Theriogenology 2021, 161, 200–209. [Google Scholar] [CrossRef]
- Food and Agriculture Organization Statistical Division (FAOSTAT) of the United Nations. Available online: https://www.fao.org/statistics/en/ (accessed on 1 July 2022).
- van Wambeke, F.; Huyghebaert, G. Current role of semen storage and artificial insemination in the turkey industry. Br. Poult. Sci. 1989, 30, 461–469. [Google Scholar] [CrossRef]
- Sexton, T.J. Oxidative and glycolytic activity of chicken and turkey spermatozoa. Comp. Biochem. Physiol. Part B Comp. Biochem. 1974, 48, 59–65. [Google Scholar] [CrossRef]
- Wishart, G.J. The effect of continuous aeration on the fertility of fowl and Turkey semen stored above 0 °C. Br. Poult. Sci. 1981, 22, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Sexton, T.J.; Giesen, A.F. Beltsville Poultry Semen Extender. Poult. Sci. 1982, 61, 1202–1208. [Google Scholar] [CrossRef]
- Marzoni, M.; Castillo, A.; Sagona, S.; Citti, L.; Rocchiccioli, S.; Romboli, I.; Felicioli, A. A proteomic approach to identify seminal plasma proteins in roosters (Gallus gallus domesticus). Anim. Reprod. Sci. 2013, 140, 216–223. [Google Scholar] [CrossRef]
- Słowińska, M.; Nynca, J.; Arnold, G.J.; Fröhlich, T.; Jankowski, J.; Kozłowski, K.; Mostek, A.; Ciereszko, A. Proteomic identification of turkey (Meleagris gallopavo) seminal plasma proteins. Poult. Sci. 2017, 96, 3422–3435. [Google Scholar] [CrossRef]
- Long, J.A. Avian Semen Cryopreservation: What Are the Biological Challenges? Poult. Sci. 2006, 85, 232–236. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.J.; O’Donovan, C.; Magrane, M.; Alpi, E.; Antunes, R.; Bely, B.; Bingley, M.; Bonilla, C.; Britto, R.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [Green Version]
- Mańkowska, A.; Brym, P.; Paukszto, Ł.; Jastrzębski, J.P.; Fraser, L. Gene polymorphisms in boar spermatozoa and their associations with post-thaw semen quality. Int. J. Mol. Sci. 2020, 21, 1902. [Google Scholar] [CrossRef] [Green Version]
- Froman, D.P.; Feltmann, A.J.; Mclean, D.J. Increased Fecundity Resulting from Semen Donor Selection Based Upon in Vitro Sperm Motility. Poult. Sci. 1997, 76, 73–77. [Google Scholar] [CrossRef]
- Froman, D.P.; Feltmann, A.J.; Rhoads, M.L.; Kirby, J.D. Sperm Mobility: A Primary Determinant of Fertility in the Domestic Fowl (Gallus domesticus). Biol. Reprod. 1999, 61, 400–405. [Google Scholar] [CrossRef] [Green Version]
- FROMAN, D.P.; McLEAN, D.J. Objective Measurement of Sperm Motility Based Upon Sperm Penetration of Accudenz®. Poult. Sci. 1996, 75, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Aire, T.A. Spermiogenesis in birds. Spermatogenesis 2014, 4, e959392. [Google Scholar] [CrossRef] [PubMed]
- Manier, M.K.; Welch, G.; Van Nispen, C.; Bakst, M.R.; Long, J. Low-mobility sperm phenotype in the domestic turkey: Impact on sperm morphometry and early embryonic death. Reprod. Domest. Anim. 2019, 54, 613–621. [Google Scholar] [CrossRef]
- Kornienko, A.E.; Guenzl, P.M.; Barlow, D.P.; Pauler, F.M. Gene regulation by the act of long non-coding RNA transcription. BMC Biol. 2013, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, A.; Tajima, A. Development and Preservation of Avian Sperm. Adv. Exp. Med. Biol. 2017, 1001, 59–73. [Google Scholar] [PubMed]
- Goley, E.D.; Welch, M.D. The ARP2/3 complex: An actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 2006, 7, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Campellone, K.G.; Welch, M.D. A nucleator arms race: Cellular control of actin assembly. Nat. Rev. Mol. Cell Biol. 2010, 11, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Dayel, M.J.; Holleran, E.A.; Mullins, R.D. Arp2/3 complex requires hydrolyzable ATP for nucleation of new actin filaments. Proc. Natl. Acad. Sci. USA 2001, 98, 14871–14876. [Google Scholar] [CrossRef] [Green Version]
- Dayel, M.J.; Mullins, R.D. Activation of Arp2/3 Complex: Addition of the First Subunit of the New Filament by a WASP Protein Triggers Rapid ATP Hydrolysis on Arp2. PLoS Biol. 2004, 2, e91. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.C.; Xu, X.-P.; Rouiller, I.; Kaksonen, M.; Sun, Y.; Belmont, L.; Volkmann, N.; Hanein, D.; Welch, M.; Drubin, D.G. Effects of Arp2 and Arp3 nucleotide-binding pocket mutations on Arp2/3 complex function. J. Cell Biol. 2005, 168, 315–328. [Google Scholar] [CrossRef]
- Robinson, R.C.; Turbedsky, K.; Kaiser, D.A.; Marchand, J.-B.; Higgs, H.N.; Choe, S.; Pollard, T.D. Crystal Structure of Arp2/3 Complex. Science 2001, 294, 1679–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Kwon, W.S.; Rahman, M.S.; Yoon, S.J.; Park, Y.J.; Pang, M.G. Actin-related protein 2/3 complex-based actin polymerization is critical for male fertility. Andrology 2015, 3, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannarella, R.; Condorelli, R.A.; Mongioì, L.M.; La Vignera, S.; Calogero, A.E. Molecular biology of spermatogenesis: Novel targets of apparently idiopathic male infertility. Int. J. Mol. Sci. 2020, 21, 1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, W.; Sun, Y.; Zeng, Z. Antimicrobial use and antimicrobial resistance in food animals. Environ. Sci. Pollut. Res. 2018, 25, 18377–18384. [Google Scholar] [CrossRef]
- Kozyreva, V.K.; McLaughlin, S.L.; Livengood, R.H.; Calkins, R.A.; Kelley, L.C.; Rajulapati, A.; Ice, R.J.; Smolkin, M.B.; Weed, S.A.; Pugacheva, E.N. NEDD9 regulates actin dynamics through cortactin deacetylation in an AURKA/HDAC6-dependent manner. Mol. Cancer Res. 2014, 12, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Yu, S.; Mipam, T.D.; Yang, F.; Zhao, W.; Liu, W.; Cao, S.Z.; Shen, L.; Zhao, F.; Sun, L.; et al. Comparative analysis of testis transcriptomes associated with male infertility in cattleyak. Theriogenology 2017, 88, 28–42. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, J.; Liu, L.; Li, X.; Liu, S.; Xia, Q.; Shi, J. Annexin A1 translocates to nucleus and promotes the expression of pro-inflammatory cytokines in a PKC-dependent manner after OGD/R. Sci. Rep. 2016, 6, 27028. [Google Scholar] [CrossRef] [Green Version]
- Munuce, M.J.; Marini, P.E.; Teijeiro, J.M. Expression profile and distribution of Annexin A1, A2 and A5 in human semen. Andrologia 2019, 51, e13224. [Google Scholar] [CrossRef]
- Sargent, K.M.; Lu, N.; Clopton, D.T.; Pohlmeier, W.E.; Brauer, V.M.; Ferrara, N.; Silversides, D.W.; Cupp, A.S. Loss of Vascular Endothelial Growth Factor A (VEGFA) Isoforms in Granulosa Cells Using pDmrt-1-Cre or Amhr2-Cre Reduces Fertility by Arresting Follicular Development and by Reducing Litter Size in Female Mice. PLoS ONE 2015, 10, e0116332. [Google Scholar] [CrossRef] [Green Version]
- Sá, R.; Miranda, C.; Carvalho, F.; Barros, A.; Sousa, M. Expression of stem cell markers: OCT4, KIT, ITGA6, and ITGB1 in the male germinal epithelium. Syst. Biol. Reprod. Med. 2013, 59, 233–243. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Xu, W.; Xia, Q.; Gu, Y.; Song, W.; Zhang, X.; Yang, Y.; Wang, W.; Li, H.; et al. Androgen promotes differentiation of PLZF+ spermatogonia pool via indirect regulatory pattern. Cell Commun. Signal. 2019, 17, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamori, N.; Tominaga, K.; Sato, T.; Riehle, K.; Iwamori, T.; Ohkawa, Y.; Coarfa, C.; Ono, E.; Matzuk, M.M. MRG15 is required for pre-mRNA splicing and spermatogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E5408–E5415. [Google Scholar] [CrossRef] [Green Version]
- Stöckl, J.B.; Schmid, N.; Flenkenthaler, F.; Drummer, C.; Behr, R.; Mayerhofer, A.; Arnold, G.J.; Fröhlich, T. Age-related alterations in the testicular proteome of a non-human primate. Cells 2021, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.E.M.; O’Bryan, M.K.; Stanton, P.G.; O’Donnell, L. The cytoskeleton in spermatogenesis. Reproduction 2019, 157, R53–R72. [Google Scholar] [CrossRef] [Green Version]
- Peruquetti, R.L.; Taboga, S.R.; Azeredo-Oliveira, M.T.V.D. Morphological Changes of Mammalian Nucleoli during Spermatogenesis and Their Possible Role in the Chromatoid Body Assembling. ISRN Cell Biol. 2012, 2012, 829854. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Ulu, A.; Chang, J.T.; Frost, J.A. Contributions of the RhoA guanine nucleotide exchange factor Net1 to polyoma middle T antigen-mediated mammary gland tumorigenesis and metastasis. Breast Cancer Res. 2018, 20, 1–16. [Google Scholar] [CrossRef]
- Gross, N.; Strillacci, M.G.; Peñagaricano, F.; Khatib, H. Characterization and functional roles of paternal RNAs in 2–4 cell bovine embryos. Sci. Rep. 2019, 9, 20347. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.; Wu, H.; Hung, R.-J.; Terman, J.R. Enhanced Production of the Mical Redox Domain for Enzymology and F-actin Disassembly Assays. Int. J. Mol. Sci. 2021, 22, 1991. [Google Scholar] [CrossRef]
- Cabodi, S.; del Pilar Camacho-Leal, M.; Di Stefano, P.; Defilippi, P. Integrin signalling adaptors: Not only figurants in the cancer story. Nat. Rev. Cancer 2010, 10, 858–870. [Google Scholar] [CrossRef]
- Chandra, V.; Huang, P.; Potluri, N.; Wu, D.; Kim, Y.; Rastinejad, F. Multidomain integration in the structure of the HNF-4α nuclear receptor complex. Nature 2013, 495, 394–398. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Li, L.; Yu, Y.; Hu, X.; Zhang, H.; Liu, R.; Wang, R. Two mutations in the axonemal dynein heavy chain gene 5 in a Chinese asthenozoospermia patient: A case report. Medicine 2020, 99, e20813. [Google Scholar] [CrossRef] [PubMed]
- Precone, V.; Cannarella, R.; Paolacci, S.; Busetto, G.M.; Beccari, T.; Stuppia, L.; Tonini, G.; Zulian, A.; Marceddu, G.; Calogero, A.E.; et al. Male Infertility Diagnosis: Improvement of Genetic Analysis Performance by the Introduction of Pre-Diagnostic Genes in a Next-Generation Sequencing Custom-Made Panel. Front. Endocrinol. 2021, 11, 605237. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.A.; Yang, Q.Z.; Duan, J.R.; Wang, Q.; Zhang, M.Y.; Zhou, Y.F.; Liu, K.; Shi, W.G. Characteristic of PGDS potential regulation role on spermatogenesis in the Chinese mitten crab Eriocheir sinensis. Gene 2014, 543, 244–252. [Google Scholar] [CrossRef]
- Pasini, M.E.; Intra, J.; Gomulski, L.M.; Calvenzani, V.; Petroni, K.; Briani, F.; Perotti, M.E. Identification and expression profiling of Ceratitis capitata genes coding for β-hexosaminidases. Gene 2011, 473, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Ewen, K.A.; Krivanek, K.M.; Clulow, J.; Kidd, G.; Ecroyd, H.; Jones, R.C. Post-testicular sperm maturation and identification of an epididymal protein in the Japanese quail (Coturnix coturnix japonica). Reproduction 2014, 147, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Ip, Y.T.; Xu, Z. Gudu, an Armadillo repeat-containing protein, is required for spermatogenesis in Drosophila. Gene 2013, 531, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Hu, X.; Li, Z.; Wang, M.; Li, S.; Wang, X.; Lin, X.; Liao, S.; Zhang, Z.; Feng, X.; et al. Transcription Factor RFX2 Is a Key Regulator of Mouse Spermiogenesis. Sci. Rep. 2016, 6, 20435. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Ke, C.C.; Chen, Y.L.; Lin, Y.H.; Yu, I.S.; Ku, W.C.; O’Bryan, M.K.; Lin, Y.H. Deficiency of the Tbc1d21 gene causes male infertility with morphological abnormalities of the sperm mitochondria and flagellum in mice. PLoS Genet. 2020, 16, e1009020. [Google Scholar] [CrossRef]
- Becker-Heck, A.; Zohn, I.E.; Okabe, N.; Pollock, A.; Lenhart, K.B.; Sullivan-Brown, J.; McSheene, J.; Loges, N.T.; Olbrich, H.; Haeffner, K.; et al. The coiled-coil domain containing protein CCDC40 is essential for motile cilia function and left-right axis formation. Nat. Genet. 2011, 43, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Antony, D.; Becker-Heck, A.; Zariwala, M.A.; Schmidts, M.; Onoufriadis, A.; Forouhan, M.; Wilson, R.; Taylor-Cox, T.; Dewar, A.; Jackson, C.; et al. Mutations in CCDC39 and CCDC40 are the Major Cause of Primary Ciliary Dyskinesia with Axonemal Disorganization and Absent Inner Dynein Arms. Hum. Mutat. 2013, 34, 462–472. [Google Scholar] [CrossRef] [Green Version]
- Sheybani, N.; Bakhtiarizadeh, M.R.; Salehi, A. An integrated analysis of mRNAs, lncRNAs, and miRNAs based on weighted gene co-expression network analysis involved in bovine endometritis. Sci. Rep. 2021, 11, 18050. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, N.; Ulitsky, I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet. 2020, 21, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Shao, D.; Xu, K.; Lu, Z.; Lu, Z.J.; Yang, Y.T.; Zhang, Q.C. RISE: A database of RNA interactome from sequencing experiments. Nucleic Acids Res. 2018, 46, D194–D201. [Google Scholar] [CrossRef] [PubMed]
- Graindorge, A.; Pinheiro, I.; Nawrocka, A.; Mallory, A.C.; Tsvetkov, P.; Gil, N.; Carolis, C.; Buchholz, F.; Ulitsky, I.; Heard, E.; et al. In-cell identification and measurement of RNA-protein interactions. Nat. Commun. 2019, 10, 5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Wang, M.; Liu, X.; Chang, G.; Chen, Y.; An, G.; Yan, L.; Gao, S.; Xu, Y.; Cui, Y.; Dong, J.; et al. Single-Cell RNA Sequencing Analysis Reveals Sequential Cell Fate Transition during Human Spermatogenesis. Cell Stem Cell 2018, 23, 599–614. [Google Scholar] [CrossRef] [Green Version]
- Słowińska, M.; Liszewska, E.; Nynca, J.; Bukowska, J.; Hejmej, A.; Bilińska, B.; Szubstarski, J.; Kozłowski, K.; Jankowski, J.; Ciereszko, A. Isolation and Characterization of an Ovoinhibitor, a Multidomain Kazal-Like Inhibitor from Turkey (Meleagris gallopavo) Seminal Plasma1. Biol. Reprod. 2014, 91, 108. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.2.1; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 20 June 2022).
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [Green Version]
- Wucher, V.; Legeai, F.; Hédan, B.; Rizk, G.; Lagoutte, L.; Leeb, T.; Jagannathan, V.; Cadieu, E.; David, A.; Lohi, H.; et al. FEELnc: A tool for long non-coding RNA annotation and its application to the dog transcriptome. Nucleic Acids Res. 2017, 45, e57. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.-P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Li, A.; Zhang, J.; Zhou, Z. PLEK: A tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 311. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Han, S.; Liang, Y.; Ma, Q.; Xu, Y.; Zhang, Y.; Du, W.; Wang, C.; Li, Y. LncFinder: An integrated platform for long non-coding RNA identification utilizing sequence intrinsic composition, structural information and physicochemical property. Brief. Bioinform. 2019, 20, 2009–2027. [Google Scholar] [CrossRef]
- Fan, X.N.; Zhang, S.W.; Zhang, S.Y.; Ni, J.J. Lncrna_mdeep: An alignment-free predictor for distinguishing long non-coding rnas from protein-coding transcripts by multimodal deep learning. Int. J. Mol. Sci. 2020, 21, 5222. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, W.; Zeng, P.; Wang, J.; Geng, B.; Yang, J.; Cui, Q. LncTar: A tool for predicting the RNA targets of long noncoding RNAs. Brief. Bioinform. 2014, 16, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Ren, S.; Lu, M.; Zhang, Y.; Zhu, D.; Zhang, X.; Li, T. Computational prediction of associations between long non-coding RNAs and proteins. BMC Genom. 2013, 14, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ahmed, M.; Kim, D.R. pcr: An R package for quality assessment, analysis and testing of qPCR data. PeerJ 2018, 6, e4473. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | E | DD | ||||
|---|---|---|---|---|---|---|
| Min/Max | Mean | Min/Max | Mean | Min/Max | Mean | |
| Raw reads | 127.9/165.8 | 146.9 | 126.4/176.9 | 152.0 | 102.6/167.7 | 145.9 |
| Trimmed reads | 110.7/140.6 | 123.8 | 96/151.9 | 127.5 | 86.2/137.6 | 121.4 |
| % of trimmed reads | 81.1/86.5 | 84.3 | 76/87.2 | 83.5 | 82/84.7 | 83.2 |
| Mapped reads | 78/100 | 87.6 | 64.8/117.3 | 93.0 | 62.3/99.9 | 84.8 |
| Uniquely mapped reads | 76.2/97.6 | 85.9 | 62.6/115.8 | 91.6 | 61.3/98.4 | 83.5 |
| % of uniquely mapped reads | 97.6/98.6 | 98.00 | 96.6/98.9 | 98.3 | 98.2/98.6 | 98.4 |
| Expressed transcripts | 59,151/73,569 | 67,594.3 | 22,109/61,868 | 47,862.8 | 33,355/47,861 | 41,412.5 |
| Expressed genes | 36,879/43,510 | 40,405.3 | 17,596/34,711 | 28,438.5 | 21,196/27,526 | 24,970.2 |
| E vs. T | DD vs. T | E vs. DD | |
|---|---|---|---|
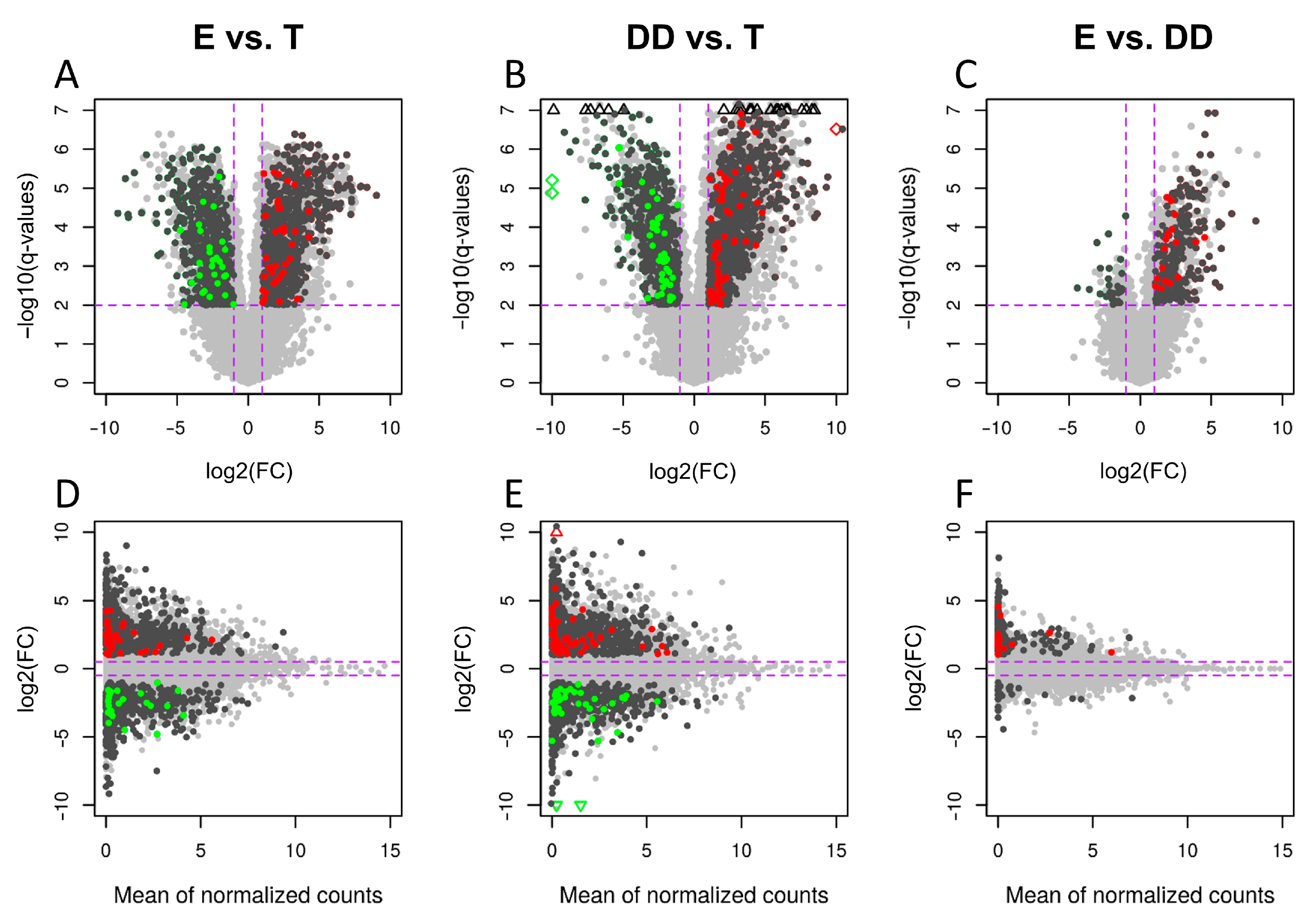
| All | 139 | 182 | 25 |
| Up-regulated | 93 | 132 | 25 |
| Down-regulated | 46 | 50 | 0 |
| Group | Process | Genes |
|---|---|---|
| T/(E + DD) | actin binding actin filament binding actin cytoskeleton organization actin filament-based process actin filament organization regulation of actin filament length regulation of protein phosphorylation apoptotic process response to oxygen-containing compound response to stress solute: cation symporter activity response to estrogen | ARPC5, CALD1, CNN3, EVL, EZR, GSN, ITGB1, MICAL2, MYH11, PARVA, POF1B, SPTBN1, TWF1, USH1C, CARMIL1, CGNL1, EPHA1, FAT1, MET, NRP1, PACSIN2, PDLIM3, PHLDB2, SDCBP, SLC9A3R1, SLIT2, SORBS1, WDR1, ANXA1, IQGAP2, ACVR1, APP, BMP7, CTNNB1, HBEGF, NEDD9, PKD2, SPRY2, THBS1, ACKR3, BAG3, GATA3, LEF1, NET1, PTPRK, ATP1A1, CAT, DCN, MMP2, GCNT1, SOD, PRDX1, ATP6V0A4, PTPPK, FIGF, MYLK, SLC15A1, SLC34A2, KRT19 |
| E/(T + DD) | calcium ion binding reproductive process lipid metabolism | CDHR2, CLGN, HPGDS, LRP2, CDHR4, PLA2G12B, CDH20, DNAH5, HEXB, HNF4A, RDH10 |
| DD/(T + E) | axonemal dynein complex assembly microtubule cytoskeleton organization microtubule-based movement inner dynein arm assembly outer dynein arm assembly flagellated sperm motility | ARMC4, CCDC40, DNAH7, LRRC6, PIH1D3, ZMYND10, DEUP1, NDC80, PLK4, RSPH9, TTC26, CFAP221, ROPN1L, CFAP69 |
| Relations/Interactions | Cis | Trans | lncRNA-mRNA | lncRNA-Protein |
|---|---|---|---|---|
| E vs. T | 0/23 | 519/15,190 | 3302/10,171 | 5845/9712 |
| DD vs. T | 0/30 | 749/22,983 | 4875/13,938 | 7335/14,172 |
| E vs. DD | 0/1 | 99/759 | 221/317 | 405/512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jastrzebski, J.P.; Lipka, A.; Majewska, M.; Makowczenko, K.G.; Paukszto, L.; Bukowska, J.; Dorocki, S.; Kozlowski, K.; Slowinska, M. In Silico Identification of lncRNAs Regulating Sperm Motility in the Turkey (Meleagris gallopavo L.). Int. J. Mol. Sci. 2022, 23, 7642. https://doi.org/10.3390/ijms23147642
Jastrzebski JP, Lipka A, Majewska M, Makowczenko KG, Paukszto L, Bukowska J, Dorocki S, Kozlowski K, Slowinska M. In Silico Identification of lncRNAs Regulating Sperm Motility in the Turkey (Meleagris gallopavo L.). International Journal of Molecular Sciences. 2022; 23(14):7642. https://doi.org/10.3390/ijms23147642
Chicago/Turabian StyleJastrzebski, Jan Pawel, Aleksandra Lipka, Marta Majewska, Karol G. Makowczenko, Lukasz Paukszto, Joanna Bukowska, Slawomir Dorocki, Krzysztof Kozlowski, and Mariola Slowinska. 2022. "In Silico Identification of lncRNAs Regulating Sperm Motility in the Turkey (Meleagris gallopavo L.)" International Journal of Molecular Sciences 23, no. 14: 7642. https://doi.org/10.3390/ijms23147642