An Updated Overview of Metabolomic Profile Changes in Chronic Obstructive Pulmonary Disease

 , ,
, ,
Abstract
:1. Introduction
2. COPD-Associated Biomarkers in Different Sample Types and Metabolic Pathway Analysis
2.1. Blood/Serum/Plasma Sample
2.1.1. Phenotypic Differentiation and Therapy of COPD
2.1.2. Diagnosis of COPD
2.1.3. Differential Diagnosis of COPD
2.2. Urine Sample
2.3. EBC
2.4. Lung Tissue Sample
2.5. Other Samples
2.6. Metabolic Pathway Analysis
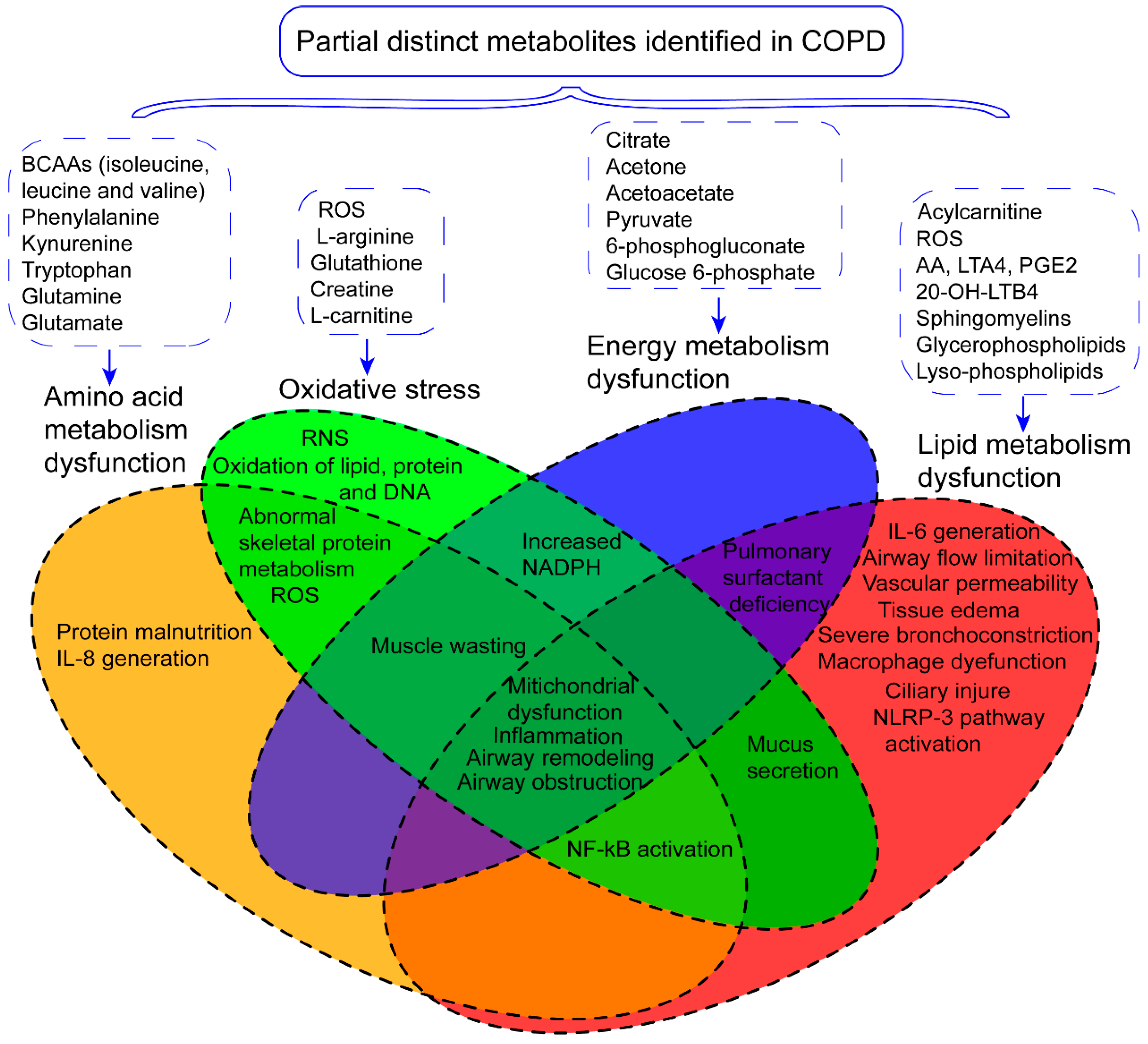
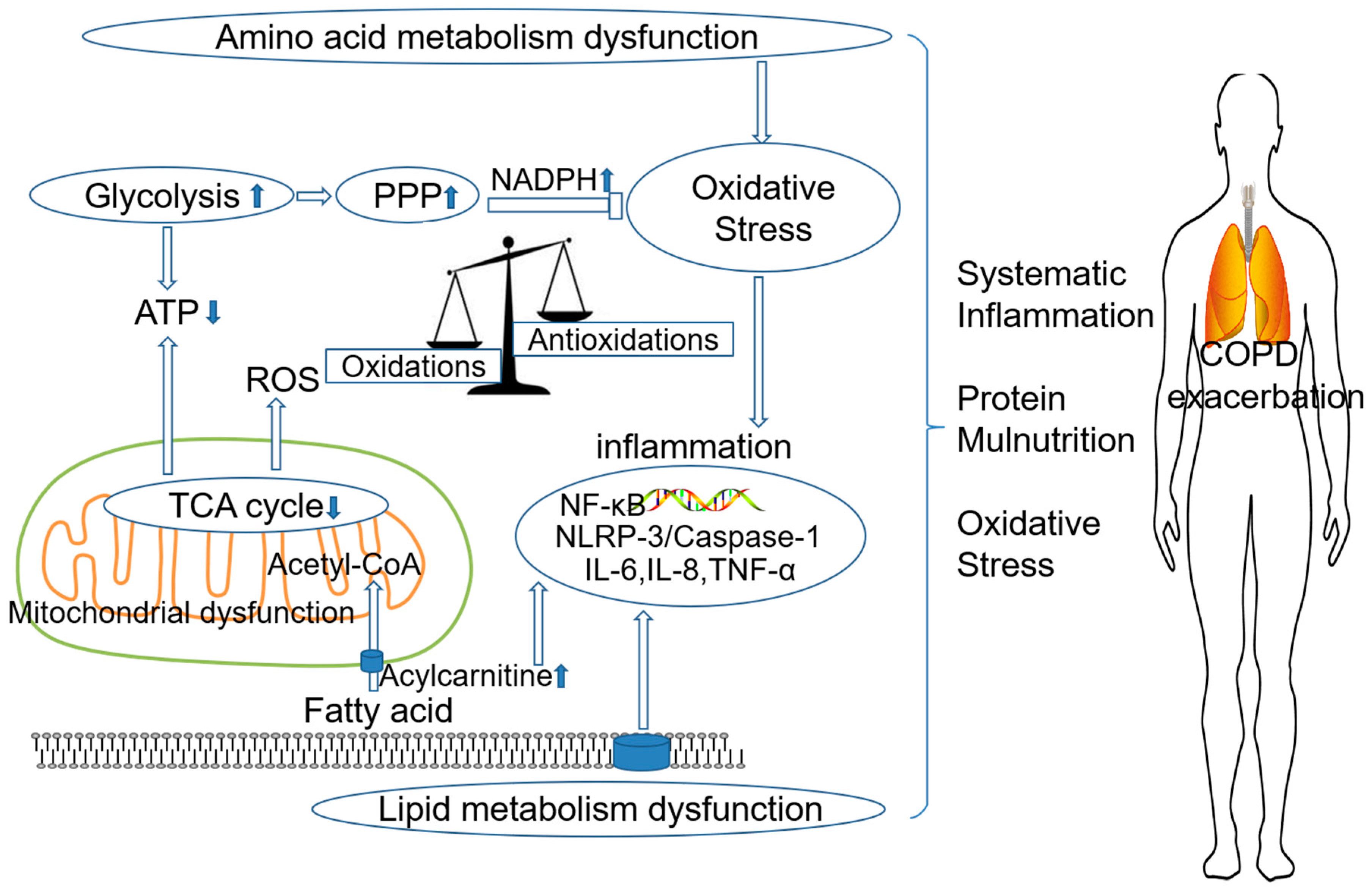
3. The Role of Metabolism Dysfunction in COPD Pathogenesis
3.1. Amino Acid Metabolism Dysfunction and COPD
3.2. Lipid Metabolism Dysfunction and COPD
3.3. Energy Metabolism Dysfunction and COPD
3.4. Oxidative Stress and COPD
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quaderi, S.A.; Hurst, J.R. The unmet global burden of COPD. Glob. Health Epidemiol. Genom. 2018, 3, e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, J.X.; Tang, D.; Zhang, J.Q.; Fang, L.Z.; Fu, W.P.; Liu, L.; Dai, L.M. Metabolic changes of different high-resolution computed tomography phenotypes of COPD after budesonide-formoterol treatment. Int. J. Chronic Obstr. Pulm. Dis. 2017, 12, 3511–3521. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, J.; Fu, X.; Ma, S.; Wang, C.; Wang, J.; Tian, S.; Liu, S.; Zhao, B.; Wang, X. LC-MS based metabolomics identification of novel biomarkers of tobacco smoke-induced chronic bronchitis. Biomed. Chromatogr. 2016, 30, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Kilk, K.; Aug, A.; Ottas, A.; Soomets, U.; Altraja, S.; Altraja, A. Phenotyping of Chronic Obstructive Pulmonary Disease Based on the Integration of Metabolomes and Clinical Characteristics. Int. J. Mol. Sci. 2018, 19, 666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Li, J.; Li, Y.; Tian, Y.; Yang, L.; Li, S. Integrating Transcriptomics, Proteomics, and Metabolomics Profiling with System Pharmacology for the Delineation of Long-Term Therapeutic Mechanisms of Bufei Jianpi Formula in Treating COPD. Biomed. Res. Int. 2017, 2017, 7091087. [Google Scholar] [CrossRef] [PubMed]
- Ubhi, B.K.; Riley, J.H.; Shaw, P.A.; Lomas, D.A.; Tal-Singer, R.; MacNee, W.; Griffin, J.L.; Connor, S.C. Metabolic profiling detects biomarkers of protein degradation in COPD patients. Eur. Respir. J. 2012, 40, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Van der Does, A.M.; Heijink, M.; Mayboroda, O.A.; Persson, L.J.; Aanerud, M.; Bakke, P.; Eagan, T.M.; Hiemstra, P.S.; Giera, M. Dynamic differences in dietary polyunsaturated fatty acid metabolism in sputum of COPD patients and controls. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Deeb, R.S.; Ma, Y.; Staudt, M.R.; Crystal, R.G.; Gross, S.S. Serum Metabolite Biomarkers Discriminate Healthy Smokers from COPD Smokers. PLoS ONE 2015, 10, e0143937. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, Y.; Liu, S.; Mao, S.; Ling, Y.; Liu, D.; He, X.; Wang, X. Metabonomic profiling of serum and urine by (1)H NMR-based spectroscopy discriminates patients with chronic obstructive pulmonary disease and healthy individuals. PLoS ONE 2013, 8, e65675. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Ma, S.; Wang, J.; Tian, S.; Fu, X.; Liu, X.; Li, Z.; Zhao, B.; Wang, X. Comparative effects of dexamethasone and bergenin on chronic bronchitis and their anti-inflammatory mechanisms based on NMR metabolomics. Mol. Biosyst. 2016, 12, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Deja, S.; Porebska, I.; Kowal, A.; Zabek, A.; Barg, W.; Pawelczyk, K.; Stanimirova, I.; Daszykowski, M.; Korzeniewska, A.; Jankowska, R.; et al. Metabolomics provide new insights on lung cancer staging and discrimination from chronic obstructive pulmonary disease. J. Pharm. Biomed. Anal. 2014, 100, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, N.; Dutta, M.; Singh, B.; Banerjee, R.; Bhattacharyya, P.; Chaudhury, K. Transcriptomics, proteomics and metabolomics driven biomarker discovery in COPD: An update. Expert Rev. Mol. Diagn. 2016, 16, 897–913. [Google Scholar] [CrossRef] [PubMed]
- Nobakht, M.G.B.F.; Aliannejad, R.; Rezaei-Tavirani, M.; Taheri, S.; Oskouie, A.A. The metabolomics of airway diseases, including COPD, asthma and cystic fibrosis. Biomarkers 2015, 20, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed]
- Bowler, R.P.; Jacobson, S.; Cruickshank, C.; Hughes, G.J.; Siska, C.; Ory, D.S.; Petrache, I.; Schaffer, J.E.; Reisdorph, N.; Kechris, K. Plasma sphingolipids associated with chronic obstructive pulmonary disease phenotypes. Am. J. Respir. Crit. Care Med. 2015, 191, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank-Quinn, C.I.; Jacobson, S.; Hughes, G.; Powell, R.L.; Petrache, I.; Kechris, K.; Bowler, R.; Reisdorph, N. Metabolomics and transcriptomics pathway approach reveals outcome-specific perturbations in COPD. Sci. Rep. 2018, 8, 17132. [Google Scholar] [CrossRef] [PubMed]
- Gulcev, M.; Reilly, C.; Griffin, T.J.; Broeckling, C.D.; Sandri, B.J.; Witthuhn, B.A.; Hodgson, S.W.; Woodruff, P.G.; Wendt, C.H. Tryptophan catabolism in acute exacerbations of chronic obstructive pulmonary disease. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 2435–2446. [Google Scholar] [CrossRef]
- Tan, L.C.; Yang, W.J.; Fu, W.P.; Su, P.; Shu, J.K.; Dai, L.M. (1)H-NMR-based metabolic profiling of healthy individuals and high-resolution CT-classified phenotypes of COPD with treatment of tiotropium bromide. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 2985–2997. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Galán, L.; del Puerto-Nevado, L.; Pérez-Rial, S.; Díaz-Gil, J.J.; González-Mangado, N.; Peces-Barba, G. Liver Growth Factor Improves Pulmonary Fibrosis Secondary to Cadmium Administration in RatsEl factor de crecimiento de hígado mejora la fibrosis pulmonar inducida tras la administración de cadmio en ratas. Archivos de Bronconeumología 2010, 46, 20–26. [Google Scholar] [CrossRef]
- Navarrete, A.; Ruperez, F.J.; Mendes, T.O.; Perez-Rial, S.; Giron-Martinez, A.; Terron-Exposito, R.; Diaz-Gil, J.J.; Peces-Barba, G.; Barbas, C.; Garcia, A. A metabolomic approach shows sphingosine 1-phosphate and lysophospholipids as mediators of the therapeutic effect of liver growth factor in emphysema. J. Pharm. Biomed. Anal. 2017, 139, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Jana, S.K.; Ghosh, N.; Das, S.K.; Joshi, M.; Bhattacharyya, P.; Chaudhury, K. Metabolomic profiling of doxycycline treatment in chronic obstructive pulmonary disease. J. Pharm. Biomed. Anal. 2017, 132, 103–108. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, F.; Pastorelli, R.; Ferrario, M.; de Blasio, F.; Marinari, S.; Brunelli, L.; Wouters, E.F.M.; Polverino, F.; Celli, B.R.; Interdisciplinary Association for Research in Lung Disease (AIMAR) Study Group. Supplementation with Qter((R)) and Creatine improves functional performance in COPD patients on long term oxygen therapy. Respir. Med. 2018, 142, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, D.A.; Alcarraz-Vizán, G.; Díaz-Moralli, S.; Reed, M.; Gómez, F.P.; Falciani, F.; Günther, U.; Roca, J.; Cascante, M. Plasma metabolic profile in COPD patients: Effects of exercise and endurance training. Metabolomics 2011, 8, 508–516. [Google Scholar] [CrossRef]
- Min, T.; Bodas, M.; Mazur, S.; Vij, N. Critical role of proteostasis-imbalance in pathogenesis of COPD and severe emphysema. J. Mol. Med. (Berl.) 2011, 89, 577–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ubhi, B.K.; Cheng, K.K.; Dong, J.; Janowitz, T.; Jodrell, D.; Tal-Singer, R.; MacNee, W.; Lomas, D.A.; Riley, J.H.; Griffin, J.L.; et al. Targeted metabolomics identifies perturbations in amino acid metabolism that sub-classify patients with COPD. Mol. Biosyst. 2012, 8, 3125–3133. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank-Quinn, C.I.; Mahaffey, S.; Justice, M.J.; Hughes, G.; Armstrong, M.; Bowler, R.P.; Reisdorph, R.; Petrache, I.; Reisdorph, N. Transient and persistent metabolomic changes in plasma following chronic cigarette smoke exposure in a mouse model. PLoS ONE 2014, 9, e101855. [Google Scholar] [CrossRef]
- Kim, V.; Davey, A.; Comellas, A.P.; Han, M.K.; Washko, G.; Martinez, C.H.; Lynch, D.; Lee, J.H.; Silverman, E.K.; Crapo, J.D.; et al. Clinical and computed tomographic predictors of chronic bronchitis in COPD: A cross sectional analysis of the COPDGene study. Respir. Res. 2014, 15, 52. [Google Scholar] [CrossRef]
- Hodgson, S.; Griffin, T.J.; Reilly, C.; Harvey, S.; Witthuhn, B.A.; Sandri, B.J.; Kunisaki, K.M.; Wendt, C.H. Plasma sphingolipids in HIV-associated chronic obstructive pulmonary disease. BMJ Open Respir. Res. 2017, 4, e000180. [Google Scholar] [CrossRef] [Green Version]
- Naz, S.; Kolmert, J.; Yang, M.; Reinke, S.N.; Kamleh, M.A.; Snowden, S.; Heyder, T.; Levanen, B.; Erle, D.J.; Skold, C.M.; et al. Metabolomics analysis identifies sex-associated metabotypes of oxidative stress and the autotaxin-lysoPA axis in COPD. Eur. Respir. J. 2017, 49, 1602322. [Google Scholar] [CrossRef]
- Novotna, B.; Abdel-Hamid, M.; Koblizek, V.; Svoboda, M.; Hejduk, K.; Rehacek, V.; Bis, J.; Salajka, F. A pilot data analysis of a metabolomic HPLC-MS/MS study of patients with COPD. Adv. Clin. Exp. Med. 2018, 27, 531–539. [Google Scholar] [CrossRef]
- Fortis, S.; Lusczek, E.R.; Weinert, C.R.; Beilman, G.J. Metabolomics in COPD Acute Respiratory Failure Requiring Noninvasive Positive Pressure Ventilation. Can. Respir. J. 2017, 2017, 9480346. [Google Scholar] [CrossRef] [PubMed]
- Cosio, B.G.; Soriano, J.B.; Lopez-Campos, J.L.; Calle-Rubio, M.; Soler-Cataluna, J.J.; de-Torres, J.P.; Marin, J.M.; Martinez-Gonzalez, C.; de Lucas, P.; Mir, I.; et al. Defining the Asthma-COPD Overlap Syndrome in a COPD Cohort. Chest 2016, 149, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Lee, Y.S.; Min, K.H.; Hur, G.Y.; Lee, S.Y.; Kang, K.H.; Rhee, C.K.; Park, S.J.; Khan, A.; Na, J.; et al. Increased urinary l-histidine in patients with asthma-COPD overlap: A pilot study. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Hu, D.; Wang, X.; Chen, Y.; Wu, Y.; Pan, L.; Li, H.; Zhang, J.; Deng, F.; Guo, X.; et al. The modification of indoor PM2.5 exposure to chronic obstructive pulmonary disease in Chinese elderly people: A meet-in-metabolite analysis. Environ. Int. 2018, 121, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Miekisch, W.; Schubert, J.K.; Noeldge-Schomburg, G.F. Diagnostic potential of breath analysis--focus on volatile organic compounds. Clin. Chim. Acta 2004, 347, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Carraro, S.; Rezzi, S.; Reniero, F.; Heberger, K.; Giordano, G.; Zanconato, S.; Guillou, C.; Baraldi, E. Metabolomics applied to exhaled breath condensate in childhood asthma. Am. J. Respir. Crit. Care Med. 2007, 175, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Craig, T.J.; Henao, M.P. Advances in managing COPD related to alpha1 -antitrypsin deficiency: An under-recognized genetic disorder. Allergy 2018, 73, 2110–2121. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, C.; Ciaramelli, C.; Fumagalli, M.; Bussei, R.; Mazzoni, V.; Viglio, S.; Iadarola, P.; Stolk, J. (1)H NMR To Explore the Metabolome of Exhaled Breath Condensate in alpha1-Antitrypsin Deficient Patients: A Pilot Study. J. Proteome Res. 2016, 15, 4569–4578. [Google Scholar] [CrossRef] [PubMed]
- De Laurentiis, G.; Paris, D.; Melck, D.; Maniscalco, M.; Marsico, S.; Corso, G.; Motta, A.; Sofia, M. Metabonomic analysis of exhaled breath condensate in adults by nuclear magnetic resonance spectroscopy. Eur. Respir. J. 2008, 32, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Lozano Sinues, P.; Meier, L.; Berchtold, C.; Ivanov, M.; Sievi, N.; Camen, G.; Kohler, M.; Zenobi, R. Breath analysis in real time by mass spectrometry in chronic obstructive pulmonary disease. Respiration 2014, 87, 301–310. [Google Scholar] [CrossRef] [PubMed]
- De Laurentiis, G.; Paris, D.; Melck, D.; Montuschi, P.; Maniscalco, M.; Bianco, A.; Sofia, M.; Motta, A. Separating smoking-related diseases using NMR-based metabolomics of exhaled breath condensate. J. Proteome Res. 2013, 12, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Miravitlles, M.; Andreu, I.; Romero, Y.; Sitjar, S.; Altes, A.; Anton, E. Difficulties in differential diagnosis of COPD and asthma in primary care. Br. J. Gen. Pract. 2012, 62, e68–e75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniscalco, M.; Paris, D.; Melck, D.J.; Molino, A.; Carone, M.; Ruggeri, P.; Caramori, G.; Motta, A. Differential diagnosis between newly diagnosed asthma and COPD using exhaled breath condensate metabolomics: A pilot study. Eur. Respir. J. 2018, 51, 1701825. [Google Scholar] [CrossRef] [PubMed]
- Montuschi, P.; Santini, G.; Mores, N.; Vignoli, A.; Macagno, F.; Shoreh, R.; Tenori, L.; Zini, G.; Fuso, L.; Mondino, C.; et al. Breathomics for Assessing the Effects of Treatment and Withdrawal With Inhaled Beclomethasone/Formoterol in Patients With COPD. Front. Pharmacol. 2018, 9, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zu, Y.; Huang, Q.; Chen, F.; Wang, G.; Lan, W.; Bai, C.; Lu, S.; Yue, Y.; Deng, F. Study on metabonomic characteristics of human lung cancer using high resolution magic-angle spinning 1H NMR spectroscopy and multivariate data analysis. Magn. Reson. Med. 2011, 66, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Matsumura, K.; Kitamura, N.; Takanami, Y.; Ito, S. Multi-omics analysis: Repeated exposure of a 3D bronchial tissue culture to whole-cigarette smoke. Toxicol. In Vitro 2019, 54, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, P.; Yang, L.; Li, Y.; Tian, Y.; Li, S.; Bai, Y. Integrating 3-omics data analyze rat lung tissue of COPD states and medical intervention by delineation of molecular and pathway alterations. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, J.; Li, Y.; Tian, Y.; Li, S.; Jiang, S.; Wang, Y.; Song, X. Identification of Metabolites and Metabolic Pathways Related to Treatment with Bufei Yishen Formula in a Rat COPD Model Using HPLC Q-TOF/MS. Evid.-Based Complementary Altern. Med. 2015, 2015, 956750. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, P.; Yang, L.; Li, Y.; Tian, Y.; Li, S. System biology analysis of long-term effect and mechanism of Bufei Yishen on COPD revealed by system pharmacology and 3-omics profiling. Sci. Rep. 2016, 6, 25492. [Google Scholar] [CrossRef] [PubMed]
- Wendt, C.H.; Nelsestuen, G.; Harvey, S.; Gulcev, M.; Stone, M.; Reilly, C. Peptides in Bronchoalveolar Lavage in Chronic Obstructive Pulmonary Disease. PLoS ONE 2016, 11, e0155724. [Google Scholar] [CrossRef] [PubMed]
- Lin, V.Y.; Fain, M.D.; Jackson, P.L.; Berryhill, T.F.; Wilson, L.S.; Mazur, M.; Barnes, S.J.; Blalock, J.E.; Raju, S.V.; Rowe, S.M. Vaporized E-Cigarette Liquids Induce Ion Transport Dysfunction in Airway Epithelia. Am. J. Respir. Cell Mol. Biol. 2018. [Google Scholar] [CrossRef]
- Aug, A.; Altraja, S.; Kilk, K.; Porosk, R.; Soomets, U.; Altraja, A. E-Cigarette Affects the Metabolome of Primary Normal Human Bronchial Epithelial Cells. PLoS ONE 2015, 10, e0142053. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics (Oxf. Engl.) 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [PubMed]
- Caramori, G.; Kirkham, P.; Barczyk, A.; Di Stefano, A.; Adcock, I. Molecular pathogenesis of cigarette smoking-induced stable COPD. Ann. N. Y. Acad. Sci. 2015, 1340, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Mizumura, K.; Cloonan, S.M.; Nakahira, K.; Bhashyam, A.R.; Cervo, M.; Kitada, T.; Glass, K.; Owen, C.A.; Mahmood, A.; Washko, G.R.; et al. Mitophagy-dependent necroptosis contributes to the pathogenesis of COPD. J. Clin. Investig. 2014, 124, 3987–4003. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Wu, Z.; Dai, Z.; Sun, K.; Wang, J.; Wu, G. Nutritional epigenetics with a focus on amino acids: Implications for the development and treatment of metabolic syndrome. J. Nutr. Biochem. 2016, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Emtner, M.; Hallin, R.; Arnardottir, R.H.; Janson, C. Effect of physical training on fat-free mass in patients with chronic obstructive pulmonary disease (COPD). Upsala J. Med. Sci. 2015, 120, 52–58. [Google Scholar] [CrossRef]
- Kemp, P.R.; Griffiths, M.; Polkey, M.I. Muscle wasting in the presence of disease, why is it so variable? Biol. Rev. Camb. Philos. Soc. 2019, 94, 1038–1055. [Google Scholar] [CrossRef] [PubMed]
- Vahid, I.; Abdolali, B.; Fatemeh, M.; Alireza, N.; Mehdi, S. The effects of branch-chain amino acids on fatigue in the athletes. Interv. Med. Appl. Sci. 2018, 10, 233–235. [Google Scholar] [CrossRef]
- Yoneda, T.; Yoshikawa, M.; Fu, A.; Tsukaguchi, K.; Okamoto, Y.; Takenaka, H. Plasma levels of amino acids and hypermetabolism in patients with chronic obstructive pulmonary disease. Nutrition 2001, 17, 95–99. [Google Scholar] [CrossRef]
- Engelen, M.P.; De Castro, C.L.; Rutten, E.P.; Wouters, E.F.; Schols, A.M.; Deutz, N.E. Enhanced anabolic response to milk protein sip feeding in elderly subjects with COPD is associated with a reduced splanchnic extraction of multiple amino acids. Clin. Nutr. 2012, 31, 616–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirnio, P.; Vayrynen, J.P.; Klintrup, K.; Makela, J.; Karhu, T.; Herzig, K.H.; Minkkinen, I.; Makinen, M.J.; Karttunen, T.J.; Tuomisto, A. Alterations in serum amino-acid profile in the progression of colorectal cancer: Associations with systemic inflammation, tumour stage and patient survival. Br. J. Cancer 2019, 120, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Maneechotesuwan, K.; Kasetsinsombat, K.; Wongkajornsilp, A.; Barnes, P.J. Decreased indoleamine 2,3-dioxygenase activity and IL-10/IL-17A ratio in patients with COPD. Thorax 2013, 68, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Maneechotesuwan, K.; Wongkajornsilp, A.; Adcock, I.M.; Barnes, P.J. Simvastatin Suppresses Airway IL-17 and Upregulates IL-10 in Patients With Stable COPD. Chest 2015, 148, 1164–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flaring, U.B.; Rooyackers, O.E.; Wernerman, J.; Hammarqvist, F. Glutamine attenuates post-traumatic glutathione depletion in human muscle. Clin. Sci. (Lond.) 2003, 104, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.P.; de Abreu, M.G.; Pelosi, P.; Rocco, P.R. Exogenous Glutamine in Respiratory Diseases: Myth or Reality? Nutrients 2016, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Sergi, G.; Coin, A.; Marin, S.; Vianello, A.; Manzan, A.; Peruzza, S.; Inelmen, E.M.; Busetto, L.; Mulone, S.; Enzi, G. Body composition and resting energy expenditure in elderly male patients with chronic obstructive pulmonary disease. Respir Med. 2006, 100, 1918–1924. [Google Scholar] [CrossRef] [PubMed]
- Rutkowsky, J.M.; Knotts, T.A.; Ono-Moore, K.D.; McCoin, C.S.; Huang, S.; Schneider, D.; Singh, S.; Adams, S.H.; Hwang, D.H. Acylcarnitines activate proinflammatory signaling pathways. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1378–E1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Knudsen, N.H.; Wang, G.; Qiu, W.; Naing, Z.Z.C.; Bai, Y.; Ai, X.; Lee, C.H.; Zhou, X. Genetic Control of Fatty Acid beta-Oxidation in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Biol. 2017, 56, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Nocito, L.; Kleckner, A.S.; Yoo, E.J.; Jones Iv, A.R.; Liesa, M.; Corkey, B.E. The extracellular redox state modulates mitochondrial function, gluconeogenesis, and glycogen synthesis in murine hepatocytes. PLoS ONE 2015, 10, e0122818. [Google Scholar] [CrossRef]
- Xu, W.F.; Upur, H.; Wu, Y.H.; Mamtimin, B.; Yang, J.; Ga, Y.J.; You, L. Metabolomic changes in patients with chronic obstructive pulmonary disease with abnormal Savda syndrome. Exp. Ther. Med. 2015, 9, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Rong, X.; Duerr, M.A.; Hermanson, D.J.; Hedde, P.N.; Wong, J.S.; Vallim, T.Q.; Cravatt, B.F.; Gratton, E.; Ford, D.A.; et al. Intestinal Phospholipid Remodeling Is Required for Dietary-Lipid Uptake and Survival on a High-Fat Diet. Cell Metab. 2016, 23, 492–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhard, W. Lung surfactant: Function and composition in the context of development and respiratory physiology. Ann. Anat. 2016, 208, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Bodas, M.; Pehote, G.; Silverberg, D.; Gulbins, E.; Vij, N. Autophagy augmentation alleviates cigarette smoke-induced CFTR-dysfunction, ceramide-accumulation and COPD-emphysema pathogenesis. Free Radic. Biol. Med. 2018, 131, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Wang, G.; Ran, N.; Lin, H.; Wang, Z.; Guan, X.; Yuan, Y.; Fang, K.; Liu, J.; Wang, F. Inhibitory Effect of Methotrexate on Rheumatoid Arthritis Inflammation and Comprehensive Metabolomics Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole Time of Flight-Mass Spectrometry (UPLC-Q/TOF-MS). Int. J. Mol. Sci. 2018, 19, 2894. [Google Scholar] [CrossRef] [PubMed]
- Haybar, H.; Shahrabi, S.; Rezaeeyan, H.; Shirzad, R.; Saki, N. Endothelial Cells: From Dysfunction Mechanism to Pharmacological Effect in Cardiovascular Disease. Cardiovasc. Toxicol. 2019, 19, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.R.; Fessler, M.B.; Gowdy, K.M. Role for phospholipid acyl chains and cholesterol in pulmonary infections and inflammation. J. Leukoc. Biol. 2016, 100, 985–997. [Google Scholar] [CrossRef]
- Lin, C.C.; Lee, I.T.; Yang, Y.L.; Lee, C.W.; Kou, Y.R.; Yang, C.M. Induction of COX-2/PGE(2)/IL-6 is crucial for cigarette smoke extract-induced airway inflammation: Role of TLR4-dependent NADPH oxidase activation. Free Radic. Biol. Med. 2010, 48, 240–254. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, P.; Hanaoka, M.; Droma, Y.; Kubo, K. Enhanced levels of prostaglandin E2 and matrix metalloproteinase-2 correlate with the severity of airflow limitation in stable COPD. Respirology 2008, 13, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Drakatos, P.; Lykouras, D.; Sampsonas, F.; Karkoulias, K.; Spiropoulos, K. Targeting leukotrienes for the treatment of COPD? Inflamm. Allergy Drug Targets 2009, 8, 297–306. [Google Scholar] [CrossRef]
- Langen, R.C.; Korn, S.H.; Wouters, E.F. ROS in the local and systemic pathogenesis of COPD. Free Radic. Biol. Med. 2003, 35, 226–235. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, W.; Mitsuyama, H.; Watanabe, M.; Oonakahara, K.; Higashimoto, I.; Osame, M.; Arimura, K. Effects of omega-3 polyunsaturated fatty acids on inflammatory markers in COPD. Chest 2005, 128, 3817–3827. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Takahashi, H.; Kasai, C.; Kiyokawa, N.; Watanabe, T.; Fujii, S.; Kashiwagura, T.; Honma, M.; Satake, M.; Shioya, T. Effects of nutritional supplementation combined with low-intensity exercise in malnourished patients with COPD. Respir. Med. 2010, 104, 1883–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natanek, S.A.; Gosker, H.R.; Slot, I.G.; Marsh, G.S.; Hopkinson, N.S.; Moxham, J.; Kemp, P.R.; Schols, A.M.; Polkey, M.I. Pathways associated with reduced quadriceps oxidative fibres and endurance in COPD. Eur. Respir. J. 2013, 41, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Michaeloudes, C.; Kirkham, P.; Adcock, I.M.; Chung, K.F. Mitochondrial reactive oxygen species and glycolysis in airway smooth muscle cell proliferation in COPD. Eur. Respir. J. 2015, 46 (Suppl. 59), OA488. [Google Scholar] [CrossRef]
- McGillick, E.V.; Morrison, J.L.; McMillen, I.C.; Orgeig, S. Intrafetal glucose infusion alters glucocorticoid signaling and reduces surfactant protein mRNA expression in the lung of the late-gestation sheep fetus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R538–R545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaeloudes, C.; Kuo, C.H.; Haji, G.; Finch, D.K.; Halayko, A.J.; Kirkham, P.; Chung, K.F.; Adcock, I.M.; COPDMAP. Metabolic re-patterning in COPD airway smooth muscle cells. Eur. Respir. J. 2017, 50, 1700202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaeloudes, C.; Bhavsar, P.K.; Mumby, S.; Chung, K.F.; Adcock, I.M. Dealing with Stress: Defective Metabolic Adaptation in Chronic Obstructive Pulmonary Disease Pathogenesis. Ann. Am. Thorac. Soc. 2017, 14, S374–S382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurfluh, S.; Nickler, M.; Ottiger, M.; Steuer, C.; Kutz, A.; Christ-Crain, M.; Zimmerli, W.; Thomann, R.; Hoess, C.; Henzen, C.; et al. Association of adrenal hormone metabolites and mortality over a 6-year follow-up in COPD patients with acute exacerbation. Clin. Chem. Lab. Med. 2018, 56, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Gosker, H.R.; Hesselink, M.K.; Duimel, H.; Ward, K.A.; Schols, A.M. Reduced mitochondrial density in the vastus lateralis muscle of patients with COPD. Eur. Respir. J. 2007, 30, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkham, P.A.; Barnes, P.J. Oxidative stress in COPD. Chest 2013, 144, 266–273. [Google Scholar] [CrossRef]
- Rahman, I. The role of oxidative stress in the pathogenesis of COPD: Implications for therapy. Treat. Respir. Med. 2005, 4, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.R.; Zhao, L.; Sancheti, H.; Sundar, I.K.; Rahman, I.; Cadenas, E. Short-term cigarette smoke exposure induces reversible changes in energy metabolism and cellular redox status independent of inflammatory responses in mouse lungs. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L889–L898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csiszar, A.; Podlutsky, A.; Wolin, M.S.; Losonczy, G.; Pacher, P.; Ungvari, Z. Oxidative stress and accelerated vascular aging: Implications for cigarette smoking. Front. Biosci. (Landmark Ed.) 2009, 14, 3128–3144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Z.; Venardos, K.; Chin-Dusting, J.; Kaye, D.M. Adverse effects of cigarette smoke on NO bioavailability: Role of arginine metabolism and oxidative stress. Hypertension 2006, 48, 278–285. [Google Scholar] [CrossRef]
- Xu, W.; Kaneko, F.T.; Zheng, S.; Comhair, S.A.; Janocha, A.J.; Goggans, T.; Thunnissen, F.B.; Farver, C.; Hazen, S.L.; Jennings, C.; et al. Increased arginase II and decreased NO synthesis in endothelial cells of patients with pulmonary arterial hypertension. FASEB J. 2004, 18, 1746–1748. [Google Scholar] [CrossRef] [Green Version]
- Dua, K.; Malyla, V.; Singhvi, G.; Wadhwa, R.; Krishna, R.V.; Shukla, S.D.; Shastri, M.D.; Chellappan, D.K.; Maurya, P.K.; Satija, S.; et al. Increasing complexity and interactions of oxidative stress in chronic respiratory diseases: An emerging need for novel drug delivery systems. Chem. Biol. Interact. 2019, 299, 168–178. [Google Scholar] [CrossRef]
- Zheng, L.; Qin, J.; Sun, L.; Gui, L.; Zhang, C.; Huang, Y.; Deng, W.; Huang, A.; Sun, D.; Luo, M. Intrahepatic upregulation of MRTF-A signaling contributes to increased hepatic vascular resistance in cirrhotic rats with portal hypertension. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 303–310. [Google Scholar] [CrossRef]
- Mohammed, A.; Gutta, V.; Ansari, M.S.; Saladi Venkata, R.; Jamil, K. Altered antioxidant enzyme activity with severity and comorbidities of chronic obstructive pulmonary disease (COPD) in South Indian population. COPD Res. Pract. 2017, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Faucher, M.; Steinberg, J.G.; Barbier, D.; Hug, F.; Jammes, Y. Influence of chronic hypoxemia on peripheral muscle function and oxidative stress in humans. Clin. Physiol. Funct. Imaging 2004, 24, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Witte, K.K.; Clark, A.L. Micronutrients and their supplementation in chronic cardiac failure. An update beyond theoretical perspectives. Heart Fail. Rev. 2006, 11, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Blanca, A.J.; Ruiz-Armenta, M.V.; Zambrano, S.; Salsoso, R.; Miguel-Carrasco, J.L.; Fortuno, A.; Revilla, E.; Mate, A.; Vazquez, C.M. Leptin Induces Oxidative Stress Through Activation of NADPH Oxidase in Renal Tubular Cells: Antioxidant Effect of L-Carnitine. J. Cell. Biochem. 2016, 117, 2281–2288. [Google Scholar] [CrossRef] [PubMed]
- Csikesz, N.G.; Gartman, E.J. New developments in the assessment of COPD: Early diagnosis is key. Int. J. Chronic Obstr. Pulm. Dis. 2014, 9, 277–286. [Google Scholar] [CrossRef]
- Nambiar, S.; Bong How, S.; Gummer, J.; Trengove, R.; Moodley, Y. Metabolomics in chronic lung diseases. Respirology 2019. [Google Scholar] [CrossRef] [PubMed]
- Oliver, F. Validated high quality automated metabolome analysis of Arabidopsis Thaliana Leaf Disks. In Concepts in Plant Metabolomics; Nikolau, B.J., Wurtele, E.S., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 1–18. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Author and Year | Subjects | Sample/Platform | Metabolites Name | |
|---|---|---|---|---|
| Increase | Decrease | |||
| Bowler, R.P. et al. (2015) [15] | 129 current and former smokers from the COPD Gene cohort | Plasma/HPLC-MS | Trihexosylceramides, Dihexosylceramides, Sulfatide d18.1. N16.0, Ganglioside GD1.d18.1. N16.0 (COPD exacerbations) | Ceramide, Sphingomyelin, Ganglioside, GM3, Sphingomyelin (Emphysema) Sphingomyelin, S1P (COPD exacerbations) |
| Kilk, K. et al. (2018) [4] | COPD patients (n = 25) and control individuals (n = 21) | Serum/HPLC-MS | LysoPC | SM, Hydroxylated SM, Arginine, Proline |
| Navarrete, A. et al. (2017) [20] | Control group and CS-exposed group without (n = 6, n = 10) or with (n = 10, n = 8) LGF treatment | Plasma/LC-QTOF-MS | Lysophosphatidylcholines, Mandelic acid, Hydroxymethylbenzoic acid (Before therapy), Sphingosine, Sphingosine 1-phosphate, Lysophospholipids (After LGF therapy) | Phenylalanine, Sphingosine 1-phosphate Sphingosine, Hydroxylysine, Dodecenoic acid, Oxo-methylthiobutanoic acid, (Before therapy) |
| Brajesh, S. et al. (2017) [21] | COPD patients receiving standard therapy (n = 40) and combination of doxycycline and standard therapy (n = 60) | Serum/NMR | Formate, Citrate, Imidazole, L-arginine (After doxycycline therapy compared with Pre-treatment group) | Lactate, Fatty acid (After doxycycline therapy compared with Pre-treatment group) |
| Ubhi, B. K. et al. (2012) [6] | Controls (n = 66) and GOLD stage II (n = 70), III (n = 64) and IV (n = 44) COPD patients | Serum/NMR LC-MS/MS | Glutamine, Phenylalanine, 3-methylhistidine, Ketone bodies | Lipoproteins, BCAAs, Glycine, Creatine, N, N-dimethylglycine |
| Ubhi, B. K. et al. (2012) [25] | GOLD IV patients (n = 30) and controls (n = 30) | Serum/LC-MS/MS | Glutamine, Aspartate, Arginine | Aminoadipate |
| Novotna, B. et al. (2018) [30] | COPD patients (n = 10) and healthy controls (n = 10) | Blood/HPLC-MS/MS | Carnitine, Phenylalanine/Tyrosine | Alanine, Phenylalanine, Pyroglutamate Free Carnitine/Acylcarnitine |
| Ren, X. et al. (2016) [10] | SD male rats include control, model, dexamethasone and bergenin groups, with 10 rats in each group | Serum/NMR | Isobutyrate, Acetone, Acetoacetate, Pyruvate, Glycine, Glycerol, Threonine (Before therapy). Glutamine, Glucose (After dexamethasone therapy). Glutamine (After bergenin therapy) | Isoleucine, Leucine, Valine, Lactate, Alanine, Proline, Glutamine, Glutamate, Creatine phosphate, Glucose, Serine (Before therapy). Pyruvate, Glycine, Threonine (After dexamethasone therapy). Pyruvate, Glycine, Threonine (After bergenin therapy) |
| Cruickshank-Quinn, C. I. et al. (2014) [26] | DBA/2J mice included controls group (n = 3), CS- exposed group for different time | Plasma/LC-MS | Homocitrulline, Arginine, Phenylacetylglycine, PI (36:2), PS (28:2), TG, Adenosine, AMP, Hypoxanthine | Glycerophospholipids, Glycerolipids, Pregnanetriol, Pentadecanoylglycine |
| Ren, X. et al. (2016) [3] | 40, 60, and 80% TS groups of rats (n = 6) and control group of rats (n = 6) | Serum/LC-MS | Lysophosphatidylethanolamine, Lysophosphatidic acid (18:1), Docosahexaenoic acid, 5-hydroxyindoleacetic acid, 5′-carboxy-γ-tocopherol | 4-imidazolone-5-propionic acid, 12-hydroxyeicosatetraenoic acid, Uridine |
| Hodgson, S. et al. (2017) [28] | HIV-associated COPD patients (n = 38), controls (n = 38) | Plasma/LC-MS/MS | Kynurenine/Tryptophan ratio, Ceramide, Fatty acids | Diacylglycero |
| De Benedetto, F. et al. (2018) [22] | 90 COPD patients received supplementation with QTer® and Creatine or placebo | Plasma/LC-MS | SM (OH) C16:1, SM C18:0 (supplementation with placebo) Lysophosphatidylcholine (Supplementation with QTer® and Creatine) | Phosphatidylcholine, Sphingomyelins (Supplementation with QTer® and Creatine) |
| Rodríguez et al. (2011) [23] | the effects of exercise on COPD patients (n = 18), healthy subjects (n = 12) | Plasma/NMR | glutamine, tyrosine, alanine, valine and isoleucine, creatine, creatinine, citrate and glucose (healthy subjects after training) | lactate, succinate and pyruvate (healthy subjects after training) lactate (COPD patients after training) |
| Wang, C. et al. (2017) [2] | Stable COPD patients with phenotype E (n = 22) and phenotype M (n = 28) | Serum/NMR | ADP, Guanosine, Choline, Glycine, Proline, Tyrosine, L-alanine, L-valine, Leucine (E and M) L-threonine, (E), Malonate (M) | Acetone, Lactate (E and M) Pyruvic acid(E), Uridine(M) |
| Tan, L.C. et al. (2018) [18] | COPD patients with phenotype E (n = 20) and phenotype M (n = 24) | Serum/NMR | Fructose, Glycine, Pyruvic acid, Pyruvate, Proline, Acetone, Lipid CH2CH2CO, Threonine, Lsopropyl alcohol (E and M), Lactate, Creatine, Citric acid, L-glutamine, Maltose Ornithine, 2-hydroxyisobutyrate, L-threonine, L-valine, Glutamic acid, β-alanine, Betaine, Cyclopentane (E); N-acetylcysteine (M) | Asparagine, pyridoxine(E) Ornithine, Guanosine, Lipoprotein(M) |
| Chen, Q. et al. (2015) [8] | Healthy smokers (n = 37), COPD smokers (n = 41) and non-smokers (n = 37) | Serum/LC-MS | Fibrinogen peptide B, Myoinositol, Dimethyluric acid, N-methylnicotinate, Cysteinsulfonic acid, Glycerophosphoinositol, Phosphatidylinositol (40:7), Creatinine | Several hydrophobic unknowns (with chromatographic retention time consistent with fatty acids and lipids) |
| Deja, S. et al. (2014) [11] | COPD patients (n = 22) and lung cancer (TNM stages I, II, III, and IV) patients (n = 77) | Serum/NMR | N-acetylated, Glycoproteins, Leucine, Lysine, Mannose, Choline, Lipid (CH3 (CH2) n) (lung cancer compared with COPD) | Acetate, Citrate, Methanol (lung cancer compared with COPD) |
| Fortis, S. et al. (2017) [31] | Stable COPD patients and acute respiratory failure patients caused by COPD exacerbation, pneumonia or heart failure | Serum Urine/NMR | Glutamine, Formate, Alanine, Proline, Histidine, Creatine, Phosphate (Serum); Cis-aconitate, Oxoglutarate (Urine, stable COPD compared with acute respiratory failure) | Mannitol, Citrate (Serum); Furoylglycine, N-oxide, Methyl-2-oxovalerate, Niacinamide Nicotinamide, (Urine, stable COPD compared with acute respiratory failure) |
| Naz, S. et al. (2017) [29] | Healthy, smokers and smokers with COPD | Serum/LC–MS | Asymmetric, Asymmetric/symmetric dimethylarginine, Fatty acid, Sphingolipid pathways (female); cAMP signaling (male) | Acetyl–ornithine/Ornithine, Arginine/(citrulline+ornithine) (female) |
| Wang, L. et al. (2013) [9] | COPD patients (n = 32) and healthy controls (n = 21) | Urine/NMR | Acetate, Ketone bodies, Pyruvate, Carnosine, M-hydroxyphenylacetate, Phenylacetyglycine, | 1-methylnicotinamide, Creatinine, Lactate |
| Huang, Q. et al. (2018) [34] | 41 Chinese elderly participants including COPD patients and their healthy spouses | Urine/ICP-MS | N-formyl-l-methionine, CPA, Decanoylcarnitine, L-histidine, Spermine, Acetylcarnosine, L-octanoylcarnitine, D-glucose (COPD) Indolelactic acid, 5-phosphoribosylamine (PM2.5 exposure) | Suberylglycine, 3-dehydrocarnitine (COPD), Uric acid, Glyceric acid, 1,3-biphosphate, Methyluric acid, 4-pyridoxic acid, Dopamine 4-sulfate, (PM2.5 exposure) |
| Airoldi, C. et al. (2016) [38] | ZZ-α1-antitrypsin deficient COPD patients (n = 11) and healthy controls (n = 11) | EBC/NMR | Acetate, 2,3-butanediol propionic acid, Lactate, Butyrate acetone, Benzoate, fatty acid, Formate, Alanine, Ethanol, Acetoin, Isopropanol acetoin, Propionate, Acetate | No report |
| Laurentiis, G. et al. (2013) [41] | Healthy smokers (n = 20), COPD smokers (n = 15), PLCH patients (n = 15) | EBC/NMR | Acetate (COPD and PLCH) 2-propanol (COPD) Isobutyrate (PLCH) | 1-methylimidazole (COPD and PLCH) Isobutyrate (COPD) 2-propanol (PLCH) |
| Ishikawa, S. et al. (2019) [46] | 3D bronchial tissue | 3D bronchial tissues/LC-MS/MS | 6-phosphogluconate, Erythrose 4-phosphate Ribose 5phosphate (R5P) | Glucose 6-phosphate, Fructose 6-phosphate, Glutathione, Oxidized glutathione, ADP, ATP |
| Li, J. et al. (2017) [47] | COPD rats treated with normal saline (n = 10) and aminophylline (APL) (n = 10) | Lung tissue/LC-QTOF/MS | LTA4, 5-HETE, 20-OH-LTB4, LXA4, PGE2 (Before ALP therapy) | LTA4, 5-HETE, LTB4, LXA4, PGE2 (After ALP therapy) |
| Yang, L. et al. (2015) [48] | SD rats included control group (n = 40), COPD group (n = 40), and BYF therapy group (n = 40) | Lung tissue/HPLC-Q-TOF/MS | Linoleic acid, Acetylcholine, Arachidonic acid, 2-methoxyestradiol 20-hydroxy-PGE2, 5-HEPE, 7-Oxo-11-dodecenoic acid, Acetyl-l-leucine (Before BYF therapy) Phenylpyruvic acid, Sphinganine PC (18:1) (After BYF therapy) | Phenylpyruvic acid, Sphinganine PC (18:1) (Before BYF therapy) Linoleic acid, Acetylcholine, Arachidonic acid, 2-methoxyestradiol 20-hydroxy-PGE2, 5-HEPE, 7-Oxo-11-dodecenoic acid, Acetyl-l-leucine (After BYF therapy) |
| Zhao, P. et al. (2017) [5] | COPD rats treated with normal saline, BJF, and aminophylline | Lung tissue/LC-MS | Glutathione (After BJF therapy) | Arachidonic acid, Linoleic acid, Glycerophospholipid (After BJF therapy) |
| van der Doesa, A. M. (2018) [7] | Smoking controls, COPD patients in a stable or acute exacerbation phase | Sputum/LC-MS | No report | ALA, EPA, LA, HEPEs, HDHAs, HETEs, LTB4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, N.; Pang, Z.; Gu, Y.; Pan, H.; Zuo, X.; Guan, X.; Yuan, Y.; Wang, Z.; Guo, Y.; Cui, Z.; et al. An Updated Overview of Metabolomic Profile Changes in Chronic Obstructive Pulmonary Disease. Metabolites 2019, 9, 111. https://doi.org/10.3390/metabo9060111
Ran N, Pang Z, Gu Y, Pan H, Zuo X, Guan X, Yuan Y, Wang Z, Guo Y, Cui Z, et al. An Updated Overview of Metabolomic Profile Changes in Chronic Obstructive Pulmonary Disease. Metabolites. 2019; 9(6):111. https://doi.org/10.3390/metabo9060111
Chicago/Turabian StyleRan, Nan, Zhiqiang Pang, Yinuo Gu, He Pan, Xu Zuo, Xuewa Guan, Yuze Yuan, Ziyan Wang, Yingqiao Guo, Zixu Cui, and et al. 2019. "An Updated Overview of Metabolomic Profile Changes in Chronic Obstructive Pulmonary Disease" Metabolites 9, no. 6: 111. https://doi.org/10.3390/metabo9060111